As an adult, you have experienced an astounding number of mutations during your lifetime – several quadrillion, to be specific. This number may seem unfathomable, but it’s true. It’s natural to wonder how you’re still alive with such an immense number of mutations. How do we even know how many mutations have occurred in our cells? And what are the consequences of all these mutations?
Let’s explore the staggering number of mutations that occur within each of us and how this relates to the survival and genetic viability of species like humans, elephants, and mice, which have thrived for thousands of generations despite experiencing countless mutations. By coming-to-grips with these vast numbers, we can gain insight into the incredible genetic resilience of living organisms.
<< A PDF version of this post can be downloaded by clicking HERE. A YouTube video based on this post is also available at the end of this article>>
A question of individual and species survival in the face of mutations
I was inspired to talk about mutations today while listening to a YouTube channel named Bread of Life. There the host asked an intuitive and common question that I would expect any of my students in genetics or evolutionary biology to ask. She observed that for her, evolution seems to work in the opposite direction of what is proposed. Specifically, if most mutations are bad, doesn’t that mean things are breaking down instead of improving? Mutations don’t seem to benefit organisms and are, on the whole, a negative occurrence.
This objection to evolution theory can be stated as follows: if most mutations are neutral or negative, and only a few are beneficial, why don’t the negative ones accumulate over time and harm a species? In other words, are we and all other species experiencing “genetic entropy”, are we all running down over time? Does this path lead us to our inevitable extinction? These questions address the fate of species in the face of the constant occurrence of new mutations introduced during the replacement of individuals in a population.
I will begin to address these questions in what might seem to be a unique way but one that I hope you will find useful and interesting. Then I will provide a more traditional response. First, I will examine the fate of mutations that occur during the lifetime of a multicellular individual, such as myself or the reader. How many mutations occur within us, and how do we survive this onslaught of mutations? While this is not a perfect analogy to mutations in populations, I believe it will help draw some useful parallels and unique perspectives to the problem of mutations.
A quadrillion mutations and counting…
Have you ever wondered how many genetic mutations you have experienced in your lifetime? It’s important to note that this question is different from how many mutations persist in your cells at present, but we’ll get to that later. To give a rough estimate, let’s consider that an adult may have around 100 trillion cells though there is some debate on this figure. Each of these cells came from a previous cell, originating from a single cell. This means that all of your cells have a common ancestor and have divided into many different lineages throughout your body. Some of your cells have undergone hundreds of divisions, while others may have undergone thousands or even hundreds of thousands leading to the cells living in you at this very moment. For example, skin and gut cells have undergone huge numbers of divisions, while progenitors to your sex cells and some brain neurons have only undergone a few.
Each time a cell divides, it must first copy its 6+ billion base pairs of DNA before creating a new cell. If you have 100 trillion cells, this represents a minimum of 100 trillion cell divisions. However, most of the cells in your body today have been replaced many times over your lifetime. In fact, some estimates suggest that we replace the equivalent of the total number of our cells every 100 days. At that rate, a 50-year-old adult may have had 5 quadrillion cells.
Each time you copy your genome, it’s estimated that you pass on an average of 1.1 to 1.5 mutations to the daughter genome, depending on the cell type (Werner et al, 2020). Let’s assume an average of 1.25 mutations per division. Even if you only had one quadrillion cells during your life, certainly an unrealistically low number, you would have experienced a minimum of 1.25 quadrillion mutations. This number doesn’t include the large number of cells that did not survive the cellular division process or died soon after due to lethal mutations. In fact, given that we experience around 375 billion mutations per day (see below), some will occur in essential genes and cause the death of the daughter cell. That cellular death is strong natural selection occurring within your body, removing the cells with the worst mutations from your total gene pool.
How many mutations each day and each year?
The number of new cells produced by an average person per day has been estimated to range from 80 to 350 billion. For the purposes of this discussion, let’s assume that the number is 300 billion, which includes cells that spontaneously abort due to lethal mutations. Assuming a rate of 1.25 mutations per cell division, this would result in 375 billion mutations per day in the new cells. Over the course of a year, this amounts to more than 130 trillion new mutations. Ultimately, this leads us back to seeing that we have experienced quadrillions of mutations over a lifetime.
Another mind-blowing way to think about these numbers is to consider that one copy of an adult’s genome contains just over 3 billion base pairs, therefore, it’s likely that nearly every possible base pair has experienced a mutation at some point in some cell during the lifetime of that adult. So, every possible variation of our genome has been generated by each person who has lived to adulthood.
How many mutations per cell in our adult bodies?
How many mutations are present in each of our adult cells? This is where we finally start to see some reasons why we aren’t all dead! While the idea of quadrillions of mutations might seem alarming, no single cell has experienced such a high number of mutations. The number of differences between an individual genome in a single cell and the first cell at conception depends heavily on the age of the individual. The older an individual is, the more cell divisions their cells have undergone, resulting in a higher likelihood of accumulated mutations.
Furthermore, the number of accumulated mutations can vary depending on the cell type. Different cell lineages have different substitution rates, which is the rate at which mutations survive from one cell division to the next, and can lead to greater accumulation of mutations in some cells. For example, the cell lineage that produces sperm and eggs has particularly low substitution rates. (See definitions of terms at the end for an explainer on the difference between mutation rate and substitution rate)
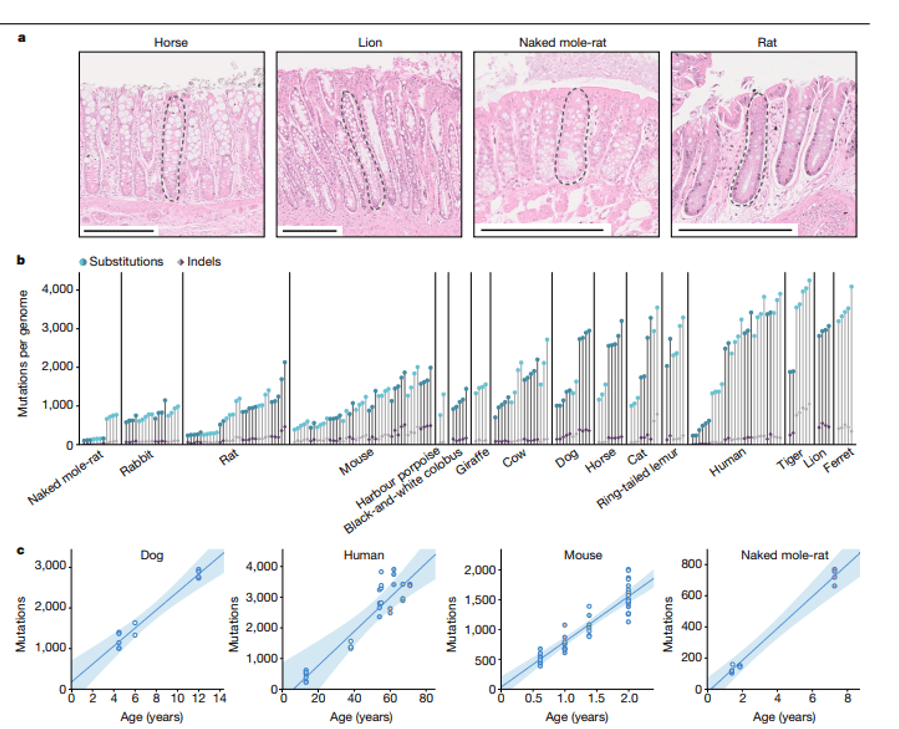
To get a handle on the number of mutations per cell, Cagan et al (2022) measured the accumulation of mutations in gut intestinal lineages (crypts) for various aged individuals and mammalian species. Their results are fascinating and illuminating, showing that mutation accumulation in intestinal cells proceeds at different rates across mammals, but they reach a similar somatic cell burden by the end of life, which is around 3000 mutations per genome. In the figure below, you can see that a 12-year-old dog has a similar number of mutations (around 2700) as a 60-year-old human. Therefore, while the substitution rate in the two organisms is different, the types of mutations and their effects are similar occurring over different time spans.
Intestinal cells replace themselves at much higher rates than other cells in the body. As a result, cancer and other genetic-based diseases are more prevalent in this tissue than in others composed of cells that have undergone fewer cell divisions and accumulated fewer mutations.
The accumulation of mutations in individual cells of our bodies does contribute to the loss of efficiency of it role is not complete loss of function. This will eventually impact the whole organisms contributing to the aging progress. While not pleasant for the individual, the impact on the species as a whole is negligible since by the time individuals accumulate this many mutations offspring will have been produced. And we will see that those offspring will inherit only a tiny fraction (100 of 10 quadrillion or 0.00000000000001%) of our mutations on the order of less than 1 of every 10 trillion mutations.
What happens to mutations in us and why don’t they do more damage than they do?
Mutations are commonly caused by errors that occur during the replication cycle of our genome. While these mutations are random, certain types of mutations are more likely to occur than others, such as C->T versus G->T. The initial mutation rate during replication can be high, but mammals have error-checking mechanisms that can scan newly formed DNA strands and fix errors, reducing the error rate to one in several million bases.
Some mutations occur in critical positions in genes, disrupting their function and causing the death of the daughter cell that receives the mutation. This is strong selection acting on those cells, resulting in many cells dying soon after they are formed, effectively eliminating the mutation from the population. Since most of the 3+ billion bases of our genome are not essential genes or genes that will ever be used in a particular tissue type, the likelihood of mutations harming functional parts of our genome is quite low. Even at 3000 mutations per cell at age 60, this only represents one mutation per every 2 million bases between our two copies of our genome in each cell. However, with millions of cells in a tissue, the likelihood that one or a few cells will be severely affected is high at that point. Which individual cells those may be can’t be predicted due to the random nature of the mutations.
What is important to note here is that the vast load of mutations is spread out among vast numbers of cells in your body. Cells that acquire mutations that significantly impact their survivorship end up dying but are replaced by cells generated by cells with less impactful mutations. Hence, the organism can survive the introduction of innumerable lethal and strongly deleterious alleles. Similarly, we observe that species can survive similar genetic loads as individuals of a species die due to lethal and strongly deleterious mutations thus removing them from the gene pool of the species.
However, mutational load matters for individuals and becomes a large part of what limits the lifespan of an organism. This leads to the question of whether species also accumulate mutations over time and experience a form of aging due to mutation pressure.
New mutations in mammalian species are abundant
Let us shift our discussion of mutations in populations of cells within an organism to populations of individuals in a species. How are new mutations introduced into populations, and how are they preserved or removed?
Let us consider the human population. First, we must determine how many new mutations are introduced annually. We can estimate this by comparing the average number of new mutations in each of us to our parents. This would be the number of mutations accumulated from conception to the formation of our father’s sperm and mother’s egg prior to our conception. Sequence comparisons of parental and offspring genomes indicates this number to be approximately 100 new mutations.
Despite each of us possessing several quadrillion mutations, we only pass on a tiny fraction (less than 0.0000001% of that number. Nonetheless, this still results in vast numbers of mutations concerning the overall population. Considering that approximately 140 million babies born each year, that is 140 million x 100 mutations or 14 billion new mutations introduced into the human population annually.
It is essential to note that this number of 140 million births per year represents only the survivors of a strong continuous natural selection occurring in utero in all mammal species. Although estimates vary, we can say that between 30 to 50% of all conceptions don’t result in live births. Therefore, a conservative estimate of the total number of human conceptions annually is 250 million.
Given 250 million conceptions and 100 new mutations found in each conception, there are 25 billion new mutations passed from sperm and egg to a potential future human population annually.
Finally, we can ask: what are the fates of these 25 billion mutations? How many will persist to shape the future of the next and subsequent generations? Put another way how many mutations will become fixed (everyone has them) in the human population resulting in population fitness decline? How are deleterious mutations removed from the population, and can they be eliminated quickly enough to prevent species degradation?
The fates of new mutations in populations
Loss of lethal or highly detrimental mutations: Up to half of those 25 billion mutations never become part of the human gene pool since 30 to 50% of conceptions result in spontaneous abortion early in embryonic development many before the mother is even aware she is pregnant either as a result of a lethal allele via new mutation directly expressed or recombination that results in the expression of previously existing lethal alleles. The latter is important because it shows if (this is a big if because it relies on continued introduction of that new mutation or increases via genetic drift which are unlikely in a large population) lethal or strongly deleterious recessive alleles increase in frequency in a population when they are not expressed (i.e.. When they are recessive but an individual carrying them still has a “normal” copy of the gene) they increase their likelihood of coming together with another lethal or strongly deleterious allele from both the sperm and egg. This then removes them both from the gene pool placing a limit on how many more of those lethal alleles can persist in a population at any one time.
If a new variant is lethal in utero than the other 99 new variants from that fertilization event will also be lost due to the death of the embryo. What we see here is that there is strong selection against any increase in the frequency of these variants (or even the existence at all copies of that variant) in the human population as they are either eliminated immediately or they are eliminated in future generations when recombination causes the alleles to be expressed.
Even if a mutation does not cause spontaneous abortion and thus lost from the population early on it may still be lost within a generation if that mutation results in conditions for the individual that make it difficult to survive until reproductive age or traits that result in reduced chances of having offspring even if reaching reproductive. It is hard to put a number on the number of variants that cause this reduced fitness but it is surely not insignificant. And remember, if deleterious allele results in an individual that does not reproduce or has reduced reproductive output then the other 99 mutations (whether they were beneficial or neutral) born with that individual will also be lost to the human population because they will not be passed to future generations.
The above situation is analogous to the figure I shared earlier showing the history of cell divisions. Many daughter cells don’t survive and are lost to the population but there are always some that survive and will become the current population of cells that make up our present-day bodies. The history of our bodies is littered with trillions of cell lines that never lived or never were able to divide again to make new cells.
So we start with 25 billion new mutations through human conception and we get down to 14 billion by the time of birth and let say another 4 billion are knocked out due to deleterious genetic disease that stop or drastically reduce the chance of reproduction. We are now at 10 billion new mutations per generation (at today’s total population size).
Many mutations are never passed to future generations due to meiosis and chance events
A large fraction of all mutations born into this world each year will never become part of subsequent generations of the gene pool. This could be because either individuals that have them never reproduce due to chance events or life-style choice or individuals who do have offspring will not pass them on because during meiotic processes in those individuals the chromosome with the new variant is not passed to their offspring.
Consider how many individuals of a particular generation either die or are disabled due to chance events prior to having an opportunity to have offspring. Their lack of offspring has nothing to do with their particular genetic variants whether they be positive, negative or neutral. But by not having offspring all the variants they carry are not carried forward to the next generation and thus do not contribute to the genetic makeup of the next gene pool.
It is generally estimated that today about 80% of women will reproduce but only 40% of males will reproduce. In the past this was probably skewed even more heavily toward female reproduction. If you consider other mammalian species male reproduction may be less than 10%. For example, think of lions or any other species in which males fight for reproductive rights of a large group of females leaving most males to never participate in the reproductive process.
Not only does this mean that only around 50% of individuals in a population reproduce resulting in a massive loss of genetic diversity each generation but that for males in particular there is an especially large bottleneck on genetic diversity running through their lineages. Genetic diversity of the Y chromosome is rapidly lost through the constant bottleneck of small numbers of males passing their Y chromosome on. Remember that even males that reproduce will only pass their Y chromosome on if they have a male offspring.
Here alone we see that of the 14 billion mutations born into the human population each year less than half will be passed to the next generation even without considering other causes of lost variation.
But that isn’t all, there is another chance event that is going to result in a massive loss of those new mutations. That event is reproduction itself! To make sperm and eggs, the genetic material has to be divided in half. This is accomplished via the process of meiosis. I didn’t say it before but of those 100 mutations you inherit from your parents you can think of them as half coming from your mother’s chromosomes in her egg and half from your father’s chromosomes in his sperm. Males have more mutations so its not exactly even but treating them equally still works for us because during meiosis we cross over our mother and fathers’ chromosomes reshuffling those variants. The point is that the sperm and the egg will only contain about half of the original 100 variants from the previous generation but also remember each will contain about 50 new mutations it has collected during its cellular divisions since conception.
Therefore, in the case of couples having one child 50% of the mutations that the parents had are lost to the next generation even as they generate new ones. If a couple had two children some more of the original variation from them will get passed on but even if they had 10 offspring because each meiotic event is independent there would still be some small percentage of genetic variation of the parents that is never passed to their offspring.
But also consider even if you have offspring the chances are that your offspring won’t have offspring. They only have a 50% chance of having offspring. Looking out over several generations we find that most human lineages terminate quite quickly. Hence the vast majority of new variants that enter into the human gene pool each don’t have much of a chance to persist over long periods of time unless they in some way provide a benefit to the individuals that have them.
Summing up this category of loss you can readily see that of the 14 billion mutations in living offspring each year far less than 50% of those will ever reach a second generation and many fewer in generations beyond just do to the realities of typical human reproduction.
But there are many other reasons why these mutations are not as harmful or as numerous as they might sound.
Many mutations create redundant variation
Many of those mutations will result in a change from one variant to another variant already present in the human population and thus is not adding a truly new variant to the human population but only represents a change from one variant to another previously existing variant. These are most often going to be examples of neutral mutations and hence have no effect on the fitness of the individual and therefore on the species as a whole.
For example, take a stretch of 100 bp of DNA sequence in between two genes and serves no regulatory role. It may not matter what any one of those bases are. It may only matter that there are 100 bases there to play structural role in how the genome is packaged for example. A mutation that changed an A to a C in the 3rd position doesn’t have any impact. This sort of variable site is common in the human genome where we see that some people have an A, some a C, some a G and the rest a T at that 3rd position. If your father had a C there but in his sperm lineage, it mutated to a T, that would be a new variant with respect to your own parental lineage but it would NOT be a new variant with respect to the human population.
As a result, many “new” mutations are nothing but changes from one variant to another variant already in existence and so don’t add to the total pool of genetic variation among humans. They do contribute to very small changes in the frequency of variants in the human gene pool.
How many of the 10 billion new mutations does this account for? I don’t know but I’ll take a stab and say tens of millions are redundant mutations.
Many mutations are neutral or very close to neutral with respect to fitness
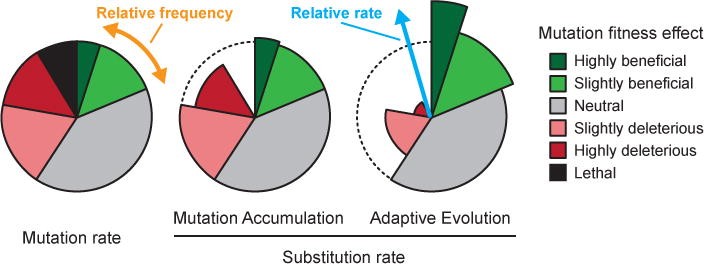
The majority of new mutations in healthy newborns are likely to be neutral or so close to neutral in terms of their effect on fitness that we call them effectively neutral. Therefore, most mutations have little to no effect on an individual’s ability to survive and reproduce, especially in large populations like that of humans. In a large population such as humans any neutral or near neutral mutation has virtually no chance of ever become fixed into the human population even if it manages to be passed down for 10,000 generations. Because selection can’t act on it, it is equally likely to become slightly less common or more common in the population. Therefore, these mutations are of minimal significance to the species as a whole. Another way of saying this is that although the mutation rates may be high the substitution rate (fixation of a new allele into the entire population) in a population is extremely low.
Some mutations are moderately deleterious
Some mutations are moderately deleterious, it is this type of mutation that some people believe are causing species, including humans, to “run down.” Another phrase often used by some people (mostly anti-evolutionists including young-earth creationists and intelligent design advocates) is to refer to genomes as subject to “genetic entropy” implying that genomes inevitably accumulate negative mutations over time. Moderately deleterious mutations are not subject to strong selection and though they are being removed via natural selection that process will take many generations to accomplish. Since new mutations are necessarily in very low frequency in large population they can be lost by accident, such as individuals failing to reproduce or meiosis failing to pass on the chromosome that contains what is a rare variant in the population.
While individual moderately or mildly deleterious mutations have no regular way of increasing their frequency in the population I need to be clear here that mutations can accumulate over time resulting in increased genetic load (see end of the this article for a detailed definition of this term) BUT accumulate in this sense means that an individual may have more deleterious mutations than their ancestor not that any individual deleterious mutation will become more common in a population. Over time larger “loads” of variants build up in the population. Again, some suggest that if this continues for many generations, it can eventually degrade the overall fitness of a species or populations within a species.
There are many reasons why this is not a necessary fate of species.
For example, consider the example above about redundant mutations. What if particular moderately or weakly negative/deleterious mutations become more common (this is a relative term, common might mean 0.0001% frequency as opposed to 0.00001%) in the population due to the same mutation occurring in multiple unrelated individuals. Even if natural selection cannot efficiently remove them from the population, there is another force that can remove deleterious mutations: mutations themselves! When the frequency of a deleterious mutation, such as a single point mutation that reduces the efficiency of an important enzyme, increases in the population, the chances that a reverse mutation will occur also increase. At some point, if an A can mutate to a C, creating a deleterious phenotype, there will be enough Cs in that position in the population that the chances of mutating the C back to an A in some sperm or eggs will occur, in which the deleterious allele is lost through mutation and replaced with a better allele. The latter mutations could be considered beneficial mutations because they restore function.
Again, I need to stress that the concern is not that a particular moderately deleterious mutation will sweep to fixation in a population and cause reduced fitness, that can’t happen,* but rather that so many deleterious mutations will occur that these will become a burden on individuals. But remember, so many of these are being removed due to lineage extinctions over generations. Natural selection need not be part of this. These mutations will be at such low frequencies that the overall burden on a particular individual having many of these is highly unlikely and has been shown not to be a problem. Just consider that we can now assess population genetic variation and mutation load from populations thousands of years ago and they were no less subject to mutation load, in fact had higher load in some cases, than populations alive today. This is not the prediction of genetic entropy which expects that past populations had lower mutation load and that it should be ever-increasing over time. In the extreme form, the original genomes were “perfect” having experienced no mutations and thus genomes can only degrade over time because of new mutations.
Some mutations are slightly, moderately, or highly beneficial.
Mutations that allow for adaptation of a population to changing environmental conditions are rare but essential. Just as natural selection works to remove any mutations that result in decreased overall fitness of an individual, it also favors mutations that provide fitness advantages over other alleles. Thus, variants with fitness advantages are expected to increase in frequency over generations, with slightly beneficial alleles increasing at a slow rate and highly beneficial alleles increasing the fastest over the same number of generations.
However, the interplay between the rate at which slightly negative alleles are removed from a population and the population’s ability to adapt to the new environment can lead to the accumulation of a genetic load of negative variants over time, relative to the number of positive mutations. But is it inevitable that populations will continuously increase the number of negative variants versus positive and neutral variants over time? Definitely not, as there are other factors to consider in addition to those mentioned above.
Populations and environments do not remain constant through time!
Some people believe that an increase in mutational load is a serious problem for species, including humans. However, these individuals, in addition to not having an appreciation for mutation/selection balance, often make the mistake of assuming that present and future conditions will be the same, or that past conditions were identical to those of today. In reality, populations are not consistently large and stable over time, and they experience varying selective pressures due to changes in the their environment which include abiotic and biotic changes.
Over thousands of generations, events will occur that significantly impact population size and introduce periods when numerous genetic variants are removed from a species. When populations are large and growing, new mutations can increase the total numbers of deleterious alleles though large population size will effectively limit each individual deleterious allele’s ability to spread through the population and become more frequent by chance. Hence, new deleterious alleles of any type will always remain very rare even as the total number of different deleterious alleles may increase and decrease over time.
However, when population size becomes small (in the case of humans it would require drastically reduced size), random events can remove variation faster than it is created by mutation. Genetic drift refers to the random fluctuations in allele frequencies that occur in small populations, especially those undergoing a bottleneck. In such populations, certain alleles may increase in frequency or disappear by chance, regardless of their fitness advantages or disadvantages. Therefore, genetic drift can counterbalance the effects of natural selection and lead to unpredictable changes in allele frequencies over time.
In the past, when populations of humans are reproductively isolated and had limited contact with each other, they effectively became smaller subpopulations for many generations. When population sizes are small, and reproductive males are even scarcer, bottleneck effects are more pronounced, resulting in the loss of genetic variation rather than an increase over time.
Returning to the effect of natural selection, large plagues throughout human history have wiped out 50% or more of the population. These events provided strong selective pressures that favored individuals with certain genotypes and hence influenced the genetic makeup of subsequent generations. Those who died carried a significant amount of genetic variation, including both deleterious and beneficial alleles, into extinction.
It is important to note that no one can assume that large selective events, such as pandemics, will not happen in the future. Similarly, disasters such as rapid climate change caused by volcanic eruptions or comet impacts can reduce human populations and split them into small subpopulations increasing the effects of genetic drift and reduce overall genetic variation in the gene pool, including the load of deleterious alleles.
Final thoughts
Genetic entropy predicts a decline in the quality and quantity of genetic information within a population, leading to a decrease in fitness and ultimately questions its survival, yet the theory lacks empirical evidence to support it. Yes, harmful mutations may accumulate in the genome over time yet there is no evidence to suggest that this will inevitably lead to the extinction of a species including the human race. While genetic mutations do occur, the idea that they inevitably lead to a loss of fitness is oversimplified and fails to take into account the complex interplay between genetic variation and natural selection.
Ironically, rather than harmful, large amounts of genetic variation are necessary for a population’s health to provide diverse phenotypes to survive events such as pandemics and climate change. More mutations due to increased births means there are also more deleterious alleles, but there are also more beneficial alleles. This is something all species experience. At any given time, some species may experience a high deleterious variant burden on their population, which could lead to, but not necessarily, reduced fitness over time. However, changing environments provide opportunities to remove large portions of those deleterious variants, increasing the species’ fitness once again. Scientists have measured populations in all phases, including decreasing, maintaining, and increasing genetic variation. In general, species with high amounts of genetic variation must have higher numbers of total deleterious alleles but are observed to have high fitness and rather than be in danger of extinction are more likely to survive than species with less genetic variation.
We have seen that as individuals we can experience far greater genetic loads than one might imagine is possible. In the end mutations will harm us as individuals but when we pass the torch of our genetic lineage to the next generation we do so in a remarkably faithful way. While very faithful we are not perfect and that is a good thing. If we made no mistakes at all future generations would be doomed to being unable to respond to changes in their living conditions.
The key for species is to find the right balance between not allowing too many mutations to pass to future generations while at the same time allowing enough mutations to allow the species to respond. It is a remarkable observation that species with different life-history traits such as growth rates, lifespans and metabolic rates have struck this balance in different ways. We saw this in the Cagan et al (2022) paper an example of this which showed that mutation rates scaled with lifespan. Canines don’t live as long as humans in part because they have a faster mutation rate. But since they replace themselves more often they can afford to make more mistakes and lose more individuals to selection against those negative alleles. But canines also are also adapting to their environment at a faster pace than do longer-lived animals. Those animals can’t afford to have such a higher mutation rate lest they create too high of a mutation load that will take too long to be eliminated from the population and therefore have a much lower mutation rate allowing for longer lifespans.

The fatal flaws of genetic entropy
Want a more direct response to the idea of genetic entropy, one that includes showing scientific data from published papers and getting into the nuts and bolts of how natural selection and mutations interact in populations? I suggest the following videos, The Fatal Flaws of Genetic Entropy, https://youtu.be/P2o_KC7sc98, The Human Genetic Load https://youtu.be/7tZScZEtp3k by Zach Hancock and also this one “5 minute myth: Genetic Entropy” by CreationMyths https://youtu.be/eb24gGAC4YE
A caveat to this presentation: I have presented several numerical values, and it is possible that you may disagree with some of my estimates. It is important to note that some of these estimates, such as the number of cells in the human body and the number of mutations, may not be entirely accurate. I have rounded off some numbers for simplicity, but I urge you not to dismiss the main points of my argument because of a slight deviation in one of the values I used. While some of my estimates could be off by a factor of ten, this would not alter the fundamental point that I am trying to make.
*There is a special situation in which a deleterious allele/mutation can increase in frequency over time in a population via natural selection. If that deleterious allele is very close to another genetic variant that has a very high fitness effect, then the beneficial allele will allow that portion of the chromosome to be selected/preserved for the next generation and so drags the deleterious allele along with it to the next generation. This will happen if the net fitness of the deleterious and beneficial allele linked together is positive. This also works for removing slightly deleterious mutations. If they are linked closely with a highly negative mutation the latter will be strongly selected against and so drag the slightly deleterious allele along with it as it is selected out of the population.
A YouTube video based on this script:
Cagan, Alex, Adrian Baez-Ortega, Natalia Brzozowska, Federico Abascal, Tim HH Coorens, Mathijs A. Sanders, Andrew RJ Lawson et al. “Somatic mutation rates scale with lifespan across mammals.” Nature 604, no. 7906 (2022): 517-524. https://www.nature.com/articles/s41586-022-04618-z
Werner, Benjamin, Jack Case, Marc J. Williams, Ketevan Chkhaidze, Daniel Temko, Javier Fernández-Mateos, George D. Cresswell et al. “Measuring single cell divisions in human tissues from multi-region sequencing data.” Nature communications 11, no. 1 (2020): 1035. https://www.nature.com/articles/s41467-020-14844-6
Definitions of terms used in this article
Mutation vs Substitution rate: Mutation rate and substitution rate are two related but distinct concepts in molecular genetic studies.
Mutation rate refers to the frequency at which new mutations arise in the DNA of a population or lineage over a given period of time. It is the rate at which changes occur in the DNA sequence of an organism, and it includes all types of mutations, whether they are beneficial, neutral, or deleterious. Mutation rate is usually expressed as the number of new mutations per site per generation or per unit of time.
Substitution rate, on the other hand, refers to the frequency at which a specific nucleotide or amino acid is replaced by another in the DNA or protein sequence of a population or lineage over a given period of time. It is the rate at which changes in the DNA or protein sequence are fixed in a population or lineage through natural selection or genetic drift. Substitution rate is usually expressed as the number of substitutions per site per unit of time.
The main difference between mutation rate and substitution rate is that mutation rate refers to the frequency of all types of mutations, while substitution rate refers specifically to the frequency of fixed mutations. In other words, while mutation rate reflects the raw material of evolution, substitution rate reflects the evolutionary process itself. Mutation rate is typically higher than substitution rate, as many mutations are not advantageous and are removed from the population by natural selection. Substitution rate, on the other hand, is influenced by both the mutation rate and the strength of natural selection or genetic drift acting on the population.
Allele: An allele is a variant form of a gene that determines a particular trait or characteristic in an organism. Genes are made up of DNA and they are responsible for encoding instructions that dictate the development, function, and behavior of cells and organisms. Alleles can differ from one another in their DNA sequence and can result in variations in traits such as eye color, blood type, or susceptibility to diseases. An organism inherits two alleles for each gene, one from each parent, and the combination of these alleles determines the genotype and phenotype of the individual.
Mutation: A mutation is a permanent alteration in the DNA sequence. Mutations can occur spontaneously or be induced by external factors such as radiation, chemicals, or viruses. Mutations can range from small changes in a single nucleotide (point mutations) to larger changes that involve the insertion, deletion, or rearrangement of entire sections of DNA. Some mutations can have no effect on an organism, while others can have harmful, beneficial, or neutral effects. Mutations can also occur in non-coding regions of DNA, which can affect gene regulation and protein production. Mutations can be inherited from one or both parents or can arise de novo in an individual’s lifetime.
Mutation load: Mutation load refers to the total burden of deleterious mutations that an individual or population carries in their DNA. Deleterious mutations are mutations that reduce an individual’s fitness or survival, such as mutations that cause diseases or impair physical or cognitive function. The mutation load can be influenced by various factors such as the rate of mutation, the size of the population, the intensity of natural selection, and the frequency of genetic recombination. In a population, the mutation load can increase over time due to the accumulation of new mutations, and this can have a negative impact on the population’s overall fitness and survival. The concept of mutation load is important in evolutionary biology and genetics as it helps to explain the effects of genetic variation on the adaptability and evolution of populations.

The Evolutionists have now become the apologeticists..
LikeLike
Great article.
“although the mutation rates may be high the substitution rate (fixation of a new allele into the entire population) in a population is extremely low.” The substitution rate per mutation is extremely low; but I thought that while the chance of a neutral mutation becoming fixed is inversely proportional to the population, the number of mutations is directly proportional, leading to the paradoxical result that provided population sizes are large enough to ignore statistical variation, the rate of fixation of new genes in a population is independent of size. Have I got this right? It’s way beyond my own intuitive understanding of statistics.
LikeLike
Mutation rate is not extremely low, as demonstrated by David Penny, Jeanson and me. It is only extremely low when we calculate rates under the evolutionary assumptions.
LikeLike
You mean substitution rates in this case.
LikeLike
Joel, when I saw you were the title of this article and use of the term “genetic entropy” I assumed you were going to address YEC claims on this topic, and especially the book entitled Genetic Entropy by John Sanford. The book is promoted by CMI and ICR, and Sanford, along with is associates Paul Price and Robert Carter, claim genetic entropy is the “Achilles heel of evolution”. I have not read Sanford’s book, but have looked over his website Geneticentropy.org. There he claims attempts to refute his thesis have failed, and under the “New Developments” he discusses various papers that supposedly confirm his genetic entropy claims, and others supposedly presently mathematical evidence against Natural Selection as a positive role in evolution. Are you familiar with Sanford’s book or the papers discussed at his website? Do you believe (as it appears) that you have effectively refuted at least Sanford’s central thesis?
More broadly, can you address the frequent claims by many YECs, including Peer Borger here, that Natural Selection has little role in evolution because it cannot create “new genetic information”? As I’ve noted before, it seems that even in the YECs own framework, it must have had a significant role in their “rapid post Flood diversification” (a.k.a hyperspeciation) model, since for most traits in most modern species, there are many more alleles than could be carried by the pairs of animals on the ark, and by any reasonable definition, new alleles constitute new genetic information.
I supposed Peer will claim, as he has often done before, that no, lots of variations and new alleles are somehow pre-programmed in the genome to appear and get passed on as needed, with little or no help from natural selection. In his past postings he has never explained exactly how this happens, and just repeatedly referred us to his book. However, today he finally emailed the chapters from his book that he promised to send. I’ll read them as I get the time, and pass along my thoughts.
By the way, a long review of Sanford’s Genetic Entropy book (paperback version) was written at Amazon by my physicist Gerald Jellison here:
Years ago when I wrote a lengthy critique of Walter Brown’s “hydroplate” flood model, Jellison helped me address the serious heat problems in the model.
Another long critique of Sanford’s book is found here:
http://blog.rongarret.info/2020/05/a-review-of-john-sanfords-genetic.html
LikeLike
Glen,
What evolutionists should do first is trying to understand our paradigm, our hypotheses. We understand yours. Yours has been refuted by observations, so better come up with something new, something testable.
This week, I have sent you several papers, which among other things, address genetic redundancy and demonstrates that natural selection does not play a role when it comes to explaining genetics at the molecular level. Genes cooperate in (scale free) networks. These networks as a whole may be subject to natural selection, but the individual nodes (genes) cooperating in these networks can be removed without problems. This is why we do not develop cancers and other diseases when some genes are removed/inactivated in regulatory networks. This is known since 30 years but largely ignored. Natural selection is maintaining the genetic status quo, just as biologists have been telling us since 1834 (!). Read my paper carefully, then you will understand why natural selection is the phlogiston of our days.
LikeLike
Peer. it is going to take me a while to wade through the writings you sent me, so be patient. As to your current comment that you understand our paradigm, clearly not. You seem to have little if any understanding of geologic and fossil record, population biology, or other disciplines highly relevant to our discussions, and instead focus entirely on genetics. I know that is your specialty, and that no one can be an expert on every field of science. However, when you don’t bother to acquire even a rudimentary understanding of other relevant diusciplines, that is a problem. It keeps you from seeing the large picture or recognizing problems in your claims and conclusions.
Joel, for example, has written extensively on the ways that microfossils in partocuilar falsify YECism and Flood Geology, while I’ve shown how tracks and other trace fossils do the same. Thousands of other workers have shown how the fossil and geologic records support an old earth and evolution in many ways. Even ICR admitted during their RATE project that the Earth’;s rocks record far too much radioactivity to be compatible with YECism (causing them to invoke multiple ad-hoc and illogical miracles to explain it). Likewise, there is extensive literature from population biology and experiments that support the role of natural selection in evolution. Have you read even a smattering of this literature? I also referred you to my collection of articles here, which summarize several lines of compelling evidence against YECIsm: http://paleo.cc/ce/compelling-evidence.htm
I don’t think you’ve ever presented any substantial counter evidence to any of this. Typically your posts just make sweeping or general statements or point us to your book.
If you want us to read and address your writings, shouldn’t you be willing to do the same? Perhaps you have some contributions to make in some aspects of genetics, but if you think your going to refute the entire evolution “paradigm” without addressing the many lines of evidence that strongly support it, good luck with that.
LikeLike
Peer, you say our paradigm has been refuted by observation. By “paradigm ” do you mean evolution? What observations refute evolution? I know of countless ones that support it (from the fossil record, population biology, and other fields) but none that refute it. In the past you’ve made comments like ‘I know my genetic findings are right so I don’t have to bother with other evidence’ (I’m paraphrasing). If other scientists took they approach why shouldn’t they say, “We already have tons of evidence that evolution is true; why should we read Peer’s book?”
As far as natural selection goes, even if some mutations are non-random as you claim, I don’t know how that rules out natural selection being involved in adaptation (why can’t it act on those mutations too?), unless you claim the non-random mutations are so intelligent or special they somehow know 1. when to appear, 2. what to consist of, and 3. how to get passed along, without natural selection being involved. Perhaps I’ll find the answer as I read more of your writings (so far I have not), but I still think it would behoove you to summarize the answers here, for those who don’t have your book, and don’t feel compelled to buy it without more incentive.
LikeLike