Young-earth creationists in the twenty-first century take a broadly inclusive view of the relationship of existing and extinct species as they relate to the kinds of organisms that God created one days four, five, and six of the creation week. For example, the image below is from a new display of the origin of the great apes at the Creation Museum and is representative of Answers in Genesis’ understanding of origin and diversification of kinds. (see also: Chimps, Orangutans, and Gorillas Evolved from a Common Ancestor on Noah’s Arkt)

The Creation Museum proposed all great apes—excluding humans—find their origin in a single proto-ape representative created by God on the sixth day of creation. Since that supernatural dust-shaping event that ape—through ordinary generation, natural selection, and mutations—has diversified both pre and post-Flood into dozens of living and extinct lineages or organisms we call species.

For young-earth creationists identifying which types of organisms were part of God’s original creation should be kind of important. It should shape how they interpret anatomical, behavioral, and genetic evidence for common ancestry and how they classify the living things in God’s creation. For example if great apes are the same kind and therefore share a common ancestor then differences in chromosome numbers, ambulatory means, metabolism, social structures and so forth require some explanation for how they could have been derived from that single common ancestor. However, if God made chimpanzees and gorillas as separate kinds they cannot share a common ancestor. Differences between them would need no explanation however, any evidence of common ancestry of chimps and gorillas would need to be explained. In the case of the great apes, the creators of the Creation Museum demonstrate that they accept the evidence for common ancestry of great apes and are striving to provide scientific explanations for the differences. For humans they emphasize the differences from great apes and seek to dismiss the evidence of common ancestry.
Despite such confident depictions of species origins, defining the meaning of the Hebrew word min translated in English as kind from scriptures has proven to be a difficult task for young earth apologists. They presume that this is the key to identifying what organisms share ancestors in the past and which are created separately during the creation week and thus do not share common ancestry. Today, many young-earth creationists ascribe to an interpretation of creation that holds that kinds are not what we recognize as species today (except possibly human beings). Rather what we identify today as species are simply the end products of processes of diversification of an originally created ancestral kind resulting in the production of many discrete morphologically and/or genetically distinguishable groups of individuals that we call species.
The broad and narrow: creationists seek broad common ancestry while also celebrating the uniqueness of species.
The move to accept large doses of common ancestry has led to some mixed messaging on the origin of kinds and the specialness of some species. For example, how might the origins pitch look different if the okapi was created separately from the savanna giraffe versus the two sharing a common giraffe-kind ancestor created by God on the sixth day?
I would submit that the answer is foundational to the apologetic message. Take the giraffe kind. Ken Ham and others have spent years telling their audience that the giraffe’s long neck is irreducibly complex. That it can’t have evolved from a shorter necked animal. The implication, sometimes directly said, is that God must have made giraffes the way they are. They are a created kind of animal. But then some visitors to the Ark Encounter must find it rather jarring when they encounter this sign explaining the origins of giraffes.

If long-necked giraffes were not found in the original creation then the long neck is not irreducibly complex after all since it was derived from a shorter neck. The appeal to a watchmaker designer as the only explanation for highly tuned and complex physiological systems that make long necks possible has far less weight once the definition of a kind is broadened to include the okapi and dozens of extinct short-necked animals as the ancestors of present day long-necked giraffe. (See Young-Earth Creationism Leads the Short-Necked Okapi to Identify as a Giraffe)
Some creationists now surmise that a shorter-necked ancestor had descendants with increasingly long necks eventually achieving the amazing necks of modern giraffes. How then are the fine-tuned mechanisms that allow long necks to exist to be explained if not by instantaneous fiat creation? They must now be explained as the end product of re-scrambled genomic variants and selection of more fit combination by the environment (ie. natural selection) to achieve characteristics not observed in the ancestors (in a very short amount of time, too). One hypothesis is that God installed the requisite genetic programming (Guliuzza, 2018) in the original short-necked ancestor at its point of creation to make such a transition possible. Nonetheless animals with shorter necks would have to transform incrementally into long-necked descendants via mechanisms generally described as ordinary biological processes rather than direct supernatural creation.
So, it seems, there has been a transition over time by the YEC camp from emphasizing evidence for God creating directly, to God creating through programming (genetics) or guiding (intervention) the development of an organism over time.
There are innumerable examples of similar incongruous messaging in the creationist’ literature. For example, I’ve written about sea otters that are said to perfectly adapted to sea life because of characteristics made directly by God, and yet the sea otter is said to be a member of a kind of animal of which all other members are adapted to land (Mixed Messages: Confusion over the Origin of Sea Otters at Answers in Genesis)
Unfortunately for creationists, defining kinds and handling the unique characteristics of descendant species doesn’t get any easier when we move our focus away from mammals.
Let’s look at an example from insects.
A case study of irreducible complexity versus post-creation hyperevolution: the bombardier beetle
Enter the bombardier beetle. Poster child for Intelligent Design and favorite cool bug example of speakers at creation conferences. Bombardier beetles have a nifty little defense system. When they are threatened, they can mix chemicals stored in an elaborate set of tubes and reservoirs in their abdomen to create a chemical reaction that results in a jet of boiling hot acid being ejected from their posterior, toward their attacker. It’s quite a feat of chemical and structural engineering.
For the creationist there can only be one answer to the origin of the bombardier beetle: God did it! To hear most young-earth apologists speak, the bombardier beetle is so specialized it’s like a mechanical pocket watch, in which each part of the mechanism is absolutely necessary and must be fully formed for the watch to have function. It seems an obvious question,but how could such an elaborate system have evolved by chance mutations and natural selection?
The creationist literature is replete with such observations. For example, here are a few quotes from Jerry Bergman on bombardier beetles:
“The entire structure involving hundreds of parts required to produce, aim and fire its poisonous mixture of unstable chemicals would be totally useless until the entire structure was completed and perfected.”
“Many different species of bombardier beetle exist, all of which are fully functional, and none of which can be used to support a Darwinian scenario. Aside from skunks (which eject a strong-smelling substance at will), no other animal has a structure even remotely similar to the bombardier beetle. If the structure had evolved through small modifications, surely many other animals would exist that likewise have evolved similar, but less (or more) complex structures. Yet, this is not the case: the bombardier beetle, although it is only one of millions of ‘unique’ animals, is completely unique in this one way.” Bergman, Jerry. 2005. Can evolution produce new organs or structures? Creation TJ 19(2) 2005.
Similarly, Melissa Webb at Answers in Genesis last year wrote: “There’s nothing like it in nature, and any sensible person knows that a tiny beetle less than an inch in size could never produce a controlled explosion by accident. It shouts an intelligent Creator.” (The Mystery of the Exploding Beetle by Melissa Webb on December 22, 2019)
So there you have it. The bombardier beetle is “completely unique” and ‘there’s nothing like it” and “no other animal has a structure even remotely similar to the bombardier beetle.” I agree that “it shouts an intelligent creator” as do all living things but do bombardier beetles necessarily shout out that that creator worked instantaneously to create these beetles without any intermediate processes?
Furthermore, just how many individuals have this ability and is it true that no other animal has a similar structure? Well, the answer for creationists is partly dependent on how they define the limits of a kind.
The rest of the story: are bombardier beetles their own kind?
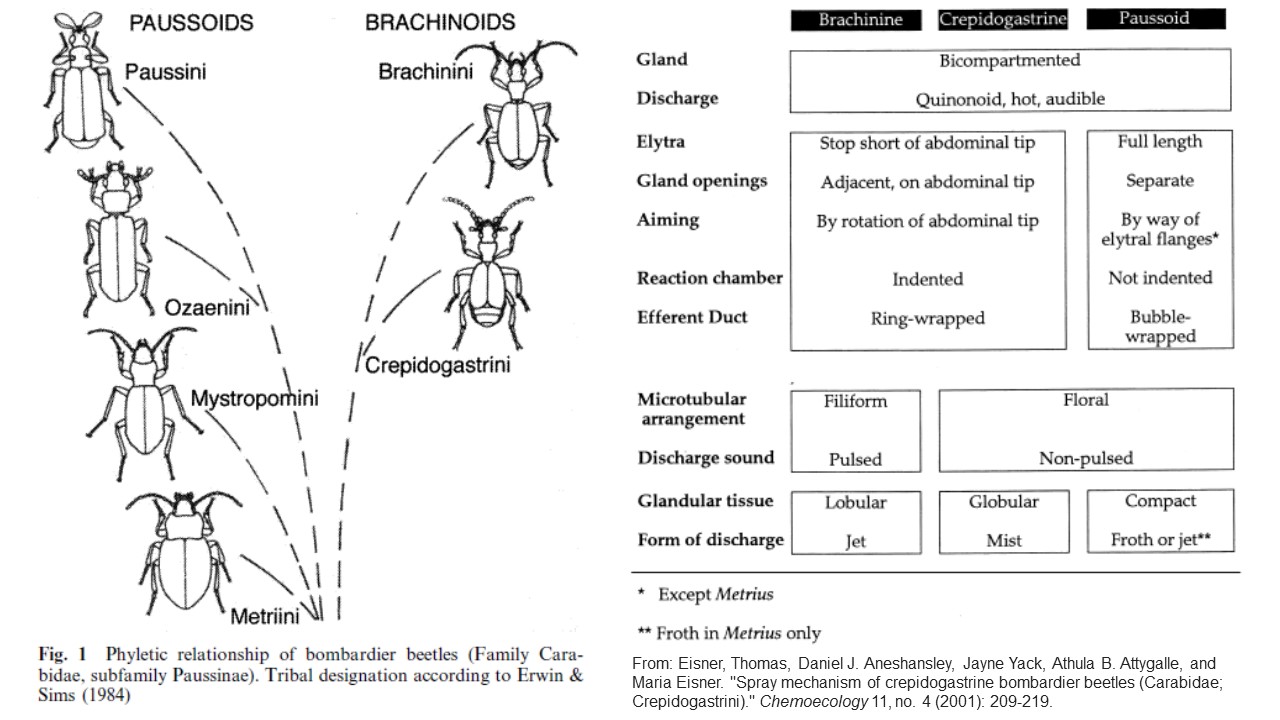
Note that above, Bergman acknowledges that “many different species of bombardier beetle exist.” Previously, having heard many young-earth speakers talk about the bombardier beetle, I inferred from them that it was either a single species or (most probably) a group of similar species all having the specialized property of spraying hot benzoquinones as a defensive mechanism. So it was surprising to learn that the term “bombardier beetle” is given to more than 500 species of beetle found in multiple genera and at least two subfamilies of a larger group, the ground beetles (family Brachinae). The bombardier beetle actually represents a diverse group of beetle species that may not even be closely related to one another. This was a shocking revelation.
Looking closer at the young-earth literature, we do find some authors who speak of the diversity of these beetles. For example, Melissa Web’s article provided a helpful graphic in which the bombardier beetles are shown divided into two varieties, the most-studied “exploding” type and those from another group of ground beetles which are sprayers, foamers, and misters (Eisner et al. 2001)of benzoquinones. Likewise, Mark Armitage (2005) reviewed the ultrastructure of the gland responsible for quinone production in one species and compared it to that of other bombardier beetle species in other genera and acknowledges there is a great variety in the structures of the glands which produce the benzoquinone among bombardier beetles. More on this later.
Why does it matter that there are over 500 species called bombardier beetles?
Let’s go back to our original question. What is a kind? It is very clear from the young-earth literature and public talks that the bombardier beetle is understood to be a uniquely created kind of organism. They claim it couldn’t have evolved from an organism that didn’t also possess the same irreducibly complex characteristics. So just what is the bombardier beetle kind and what functions / features does it include?
Beetle experts have identified over 500 species of beetles that are given the common name, bombardier beetle. This name is not a formal taxonomic designation like family, genus or species. Rather it groups all beetles based on a single distinctive character: their particular defensive behavior that includes the ejection of a noxious chemical spray via an exothermic chemical reaction.
The 500 species are not all placed in the same formal taxonomic group but rather are found in four different subfamilies called tribes (Brachinini, Paussini, Ozaenini, or Metriini) of the ground beetle family Carabidae which contains more than 36,000 species. Could the Carabidae be a created kind? If this is so, the species that possess bombardier capabilities are but a small subset of this group and suggest that this specialized defensive capability evolved from ancestors that didn’t possess this specific defensive system.

So how should we define the bombardier beetle kind?: 1) Coleoptera – all beetles (400,000+ species) are a single created kind, 2) all beetles of the family Carabidae (36,000 species) are a kind, 3) all beetles with the bombardier beetle characteristic are a single kind, or 4) there are multiple beetle kinds that share the bombardier characteristic, but were created separately.
Creationists usually distinguish kinds based upon some evidence of hybridization among species. Lacking such evidence due to a dearth of hybridization studies (as is in this case with beetles) they would likely group species by morphological similarities. Given that ground beetles share so many characteristics it is understandable that all bombardier beetles are placed within the ground beetle family (Carabidae). The modern creationist inclination is to accept all species grouped at the familial level by taxonomists as a kind. If they followed this practice for insects, they would lump the bombardier beetles into the ground beetle family despite the specialized bombardier abilities of just some of these ground beetles. Hence, the 36,000 species of ground beetles alive today must have derived from a common ancestor God created just 6000 years ago.
But a greater understanding of the limits of a kind has consequences. It invokes massive and rapid adaptive evolution within the kind and it strains what has been an effective apologetic tool to creationist speakers: the appeal to irreducible complexity as proof that God created them as they are.
A ground beetle created kind? This creates more problems than solutions for young earth creationists
If creationists place bombardier beetles in this family, what do they do with the 35,000+ species that don’t have the same irreducible and supposedly utterly unique features that bombardier beetles have?
Let’s look back at Jerry Bergman’s quote: “Many different species of bombardier beetle exist, all of which are fully functional, and none of which can be used to support a Darwinian scenario. Aside from skunks (which eject a strong-smelling substance at will), no other animal has a structure even remotely similar to the bombardier beetle. If the structure had evolved through small modifications, surely many other animals would exist that likewise have evolved similar, but less (or more) complex structures. Yet, this is not the case: the bombardier beetle, although it is only one of millions of ‘unique’ animals, is completely unique in this one way.” Bergman, Jerry. 2005. Can evolution produce new organs or structures? Creation TJ 19(2) 2005.
If Bergman is correct then bombardier beetles are unique and had to have been created directly by God. It seems like he would think that other ground beetles must be separately created and thus in a different kind. But much of what he says here is just wrong. Most importantly, the complex gland responsible for producing, storing and mixing the chemicals that are ejected from bombardier beetles is not unique.
All members of the adephagan suborder of beetles (40,000 species) including the carabids with 36,000 species and water beetles, have paired pygidial glands located in the abdomen, which are used to produce, store, mix and excrete chemicals from the beetles. They can either secrete them by oozing, spraying or crepitation (explosive spraying or misting). Within the carabid beetles there dozens of different forms of acids, ketones, phenols, quinones (the bombardier beetles) that are ejected by these glands for more than just defensive purposes.

Do young-earth apologists know about members of the ground beetle genus Galerita? These are called the false bombardier beetles because they have a similar appearance and also spray chemicals at their attacker. However, that spray is formic acid (the same substance that ants make) rather than benzoquinones (Rossini 1997). Are these also irreducibly complex and thus a unique creation and their own kind as well?
So all carabid beetles have this chemical production capacity gland but they deploy somewhat different chemicals and for different purposes. Bergman and other creationists imply—possibly due to ignorance—that the apparatus in bombardier beetles is unique when it is not. Oddly, Armitage and Mullisen (2003) mention Galerita in their paper about pygidial glands of Bombardier beetles but only mention that Galerita beetles are inhibited by the bombardier beetle spray not that these false bombardier beetles can spray them back with a different chemical. Yes, the particular chemical concoction they produce in this apparatus is different from (and more explosive than) other ground beetles, but the essential components for such a system are all present in Galerita and thousands of ground beetles of this family.
Another specialization among ground beetles as amazing as the bombardier defense
I saved the biggest and most interesting revelation for the end. For creationists that insist that bombardier beetles are specially designed, their own logic should lead them to realize that bombardier beetles cannot be a single created kind. There must be at least two created kinds of bombardier beetles, albeit with similar defensive mechanisms.
How can I say this? Looking at the differences between bombardier beetles (figure below), it becomes apparent that beetles in the subfamilies Brachinini and Paussini (and possibly others) cannot be the same kind. Young-earthers who insist that a kind is defined by the presence of supposedly irreducibly complex characteristics run into a problem, because the Paussini ground beetles are themselves highly specialized with a completely different set of unique characteristics. This is because Paussini ground beetles are also ant nest beetles.
Ant nest beetles make their home in ant nests (myrmecophiles). Yes, hundreds of Paussini bombardier beetle species live in ant nests where they are parasites on their hosts. How do they live with the ants without the ants attacking them? They excrete chemicals that identify themselves as ants. In addition, they have anatomical structure that emits sounds (Di Giulio et al, 2014, Giulio et al. 2015) that mimic ant queens causing the ants to treat them with great care. I have no doubt that if a creationist read about this, they would marvel at the amazing complexity of the auditory devices and their ability to manipulate the behavior of the ants and shout: God did it! They would consider the chemical cues and sound-making capacity as irreducibly complex characteristics that could not have originated via any natural process.
The Paussini bombardier beetles have so many unique characteristics of their own. Applying the logic found in popular creationist’ literature, we would expect them to conclude that these beetles are a separate creation—their own kind.

A narrow view of kinds focused on unique differences or a broad view of kinds focused on shared similarities?
The conundrum for the young-earth taxonomists is apparent. The ecology of living with ants as if they were ants sets the Paussini beetle apart from other beetles. At the same time, they also share so many features with other ground beetles that it is tempting to consider all ground beetles as a single kind.
To this point, young-earth popularizers lump the bombardier beetles together in a single kind. What I am suggesting is that unless they wish to go further and place them into a broader kind including all ground beetles they should realize that they may need to split the bombardier beetles into multiple created kinds. Yes, the Paussini and Brachinini bombardier beetles all excrete the same benzoquinone hot chemicals. But digging a little deeper in the literature reveals that, although they produce the same benzoquinone chemical, how they make that chemical and how they eject it from their bodies (some spray and some foam) represents some significant differences (Armitage and Mullisen, 2003; Di Guilio et al. 2015, Eisner et al. 2001; Muzzi et al, 2019a, 2019b, and Rork et al 2019) Further, the auditory and other ant-manipulation characteristics of the ant-nest beetles suggest that they deserve to be considered their own created kind.
So, what is a kind of beetle? You may be confused, and I am too. One thing is clear, the boundaries of a kind depend upon what characteristic you focus. The Creation Museum has elected to focus on the similarities of great apes versus their differences, will they likewise ignore the unique characteristics of bombardier beetles and embrace the similarities of all ground beetles or maybe even all beetles?
Post-creation rapid evolution of ground beetles?
The very fact that even a few dozen of the 500+ species of bombardier beetles possess significant anatomical differences suggests that if they truly are all one “kind” of organism then dramatic changes have occurred to the defensive systems of these beetles. The implications of this fact are that they must be far more malleable by evolutionary mechanisms than are generally recognized by young-earth and intelligent design advocates. Imagine if they were to examine a dozen other parts of the beetle anatomy such as the stribulatory—a sound-emitting—organ of the ant nest beetles? What might they conclude about the intrabaraminic (within a kind) diversity is possible? And, if this kind of diversity is, in fact, possible, what does this say about the concept of irreducible complexity?
What about taking that broader understanding of an insect kind—the direction that young-earthers have been moving in recent decades? If they are consistent with the family designation as the most likely boundary of a kind, then they are going to need to lump all ground beetles (family Carabidae) together as a single type of created life. In this scenario all ground beetles are related by ancestry to one another. If this is true, the specializations such as making benzoquinones or chemicals that mimic ant pheromones or the ability to make sounds that appeal to ants are uniquely derived traits within that ground beetle kind. Rather than being irreducibly complex, they would have to be viewed as being the products of rapid evolution of the originally created beetle kind. This rapid speciation results in a diversity of ground beetles, some of which we are awed by their incredible uniqueness.
As we have seen in this brief review, there is much more diversity of structure, function, and behavioral traits in and among these beetles than young-earth apologists either know about or recognize. They must decide if the uniqueness of bombardier beetles warrants their claim of having a special place as a unique creation or if they should come to understand the bombardier capacity as one of many evolved traits within a kind.
The short summary
- Bombardier beetles have been a favorite example of possessing a complex trait that could not have evolved and therefore must have been created by God, fully formed, rather than having evolved from non-bombardier ancestors
- There are hundreds of beetles with bombardier abilities however they don’t all perform the action in the same way suggesting at least some diversification (evolution) of their internal structures and behaviors since their creation.
- Bombardier beetles aren’t all closely related to each other. Rather some bombardier beetles–such as the ant nest beetles–are more closely related to non-bombardier beetles in other subfamilies than they are to other bombardier beetles (i.e. they are a paraphyletic group.) This is why they are placed in disparate subfamilies of a much larger one, the ground beetle family. This observation that there are at least two separate lineages of bombardier beetles is devastating to their claim of irreducible complexity and this issue hasn’t been addressed by the young-earth literature.
- All ground beetles have the same organ system referred to in this post—the pygidial gland—that bombardier beetles have. They don’t all make benzoquinones but are capable of making a range of other caustic compounds used for multiple purposes including defense.
- The presence of internal structures capable of producing and secreting chemicals from the abdomen is not special to bombardier beetles and appears to be a general property suited for adaptation into more specialized versions from a more general ancestor.
- If all ground beetles are of the same kind, one is at a loss to explain how functions such as benzoquinone foams and queen ant mimicry, thought previously by creationists to clearly be irreducibly complex, went on instead to evolve from common ancestors in just a few thousand years.
Conclusion: The broad definition of kind that is popular among creationist today contradicts 40 years of messages conveyed about bombardier beetles as unique creations that defy any evolutionary explanation. The push to explain species’ diversity as the product of rapid diversification/evolution from a created ancestral kind over just a few thousand years undermines young-earth apologists who point to specialized characteristics (the irreducible complexity claim) of individual species as incapable of having formed by known evolutionary processes.
Next up..
I want to explore the young-earth understanding of kinds of insects further by looking at other insect groups such as dragonflies, cockroaches and termites.
Update: Below is a video I made about YEC and walking stick insect origins. I called this “Kind of Confusing Part II” because it follows up on this post about bombardier beetles.
—————————————————————————————————————————
PS The young-earth literature has some catch up work to do. There have been many important studies published on bombardier beetles in the last decade that haven’t been, as yet, accounted for.. Most young-earth articles simply repeat the same story they have for 30 years and seem to be unaware–as I was until I looked–that the bombardier story is far more complex than the simple irreducible complexity they are fond of recounting.
Mark Armitage in 2003 suggested he would engage in future work to elucidate the variation among these beetles but so far none of that work has been published that I am aware of. There is an initiative at Liberty University to use bombardier beetles in biomimicry research but again, nothing new seems to have come from that yet. I have yet to find a serious discussion in the young-earth literature about the meaning of variation within the bombardier beetles or how these beetles may relate to other ground beetles. They seem content to not peek behind the curtain and find out more about a favorite example of design lest they discover things that make the story more messy.
Literature cited and used in preparation for writing this article:
Armitage, M.H. and Mullisen, L., 2003. Preliminary observations of the pygidial gland of the Bombardier Beetle, Brachinus sp., Journal of Creation 17(1):95–102.
Attygalle, Athula B., Sihang Xu, Wendy Moore, Reilly McManus, Aman Gill, and Kipling Will. “Biosynthetic origin of benzoquinones in the explosive discharge of the bombardier beetle Brachinus elongatulus.” The Science of Nature 107, no. 4 (2020): 1-11.\
Carcamo-Noriega, Edson Norberto, Shyam Sathyamoorthi, Shibdas Banerjee, Elumalai Gnanamani, Monserrat Mendoza-Trujillo, Dulce Mata-Espinosa, Rogelio Hernández-Pando, José Ignacio Veytia-Bucheli, Lourival D. Possani, and Richard N. Zare. “1, 4-Benzoquinone antimicrobial agents against Staphylococcus aureus and Mycobacterium tuberculosis derived from scorpion venom.” Proceedings of the National Academy of Sciences 116, no. 26 (2019): 12642-12647.
Di Giulio, Andrea, Simone Fattorini, Wendy Moore, James Robertson, and Emanuela Maurizi. “Form, function and evolutionary significance of stridulatory organs in ant nest beetles (Coleoptera: Carabidae: Paussini).” European Journal of Entomology 111, no. 5 (2014): 692.
Di Giulio, Andrea, Emanuela Maurizi, Francesca Barbero, Marco Sala, Simone Fattorini, Emilio Balletto, and Simona Bonelli. “The pied piper: a parasitic beetle’s melodies modulate ant behaviours.” PLoS One 10, no. 7 (2015): e0130541. https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0130541
Di Giulio, Andrea, Maurizio Muzzi, and Roberto Romani. “Functional anatomy of the explosive defensive system of bombardier beetles (Coleoptera, Carabidae, Brachininae).” Arthropod Structure & Development 44, no. 5 (2015): 468-490. https://www.sciencedirect.com/science/article/abs/pii/S1467803915000766
Eisner, Thomas, Daniel J. Aneshansley, Jayne Yack, Athula B. Attygalle, and Maria Eisner. “Spray mechanism of crepidogastrine bombardier beetles (Carabidae; Crepidogastrini).” Chemoecology 11, no. 4 (2001): 209-219.
Giglio, Anita, Pietro Brandmayr, Federica Talarico, and Tullia Zetto Brandmayr. “Current knowledge on exocrine glands in carabid beetles: structure, function and chemical compounds.” ZooKeys 100 (2011): 193.
Randy J. Guliuzza, P.E., M.D. 2018. Engineered Adaptability: Designed Mechanisms Best Explain Convergent Traits. Acts & Facts. 47 (5).
Housecroft, Catherine E. “Nature’s Chemical Weapons: Beetle Defenses.” CHIMIA International Journal for Chemistry 73, no. 5 (2019): 420-421.
Moore, Wendy. “Phylogeny of the western Hemisphere Ozaenini (Coleoptera: Carabidae: Paussinae) based on DNA sequence data.” Annals of Carnegie Museum 77, no. 1 (2008): 79-92.
Muzzi, Maurizio, Wendy Moore, and Andrea Di Giulio. “Morpho-functional analysis of the explosive defensive system of basal bombardier beetles (Carabidae: Paussinae: Metriini).” Micron 119 (2019): 24-38.
Muzzi, Maurizio, and Andrea Di Giulio. “The ant nest “bomber”: Explosive defensive system of the flanged bombardier beetle Paussus favieri (Coleoptera, Carabidae).” Arthropod structure & development 50 (2019): 24-42.
Ober, Karen A. “Phylogenetic relationships of the carabid subfamily Harpalinae (Coleoptera) based on molecular sequence data.” Molecular Phylogenetics and Evolution 24, no. 2 (2002): 228-248.
Rork, Adam M., István Mikó, and Tanya Renner. “Pygidial glands of Harpalus pensylvanicus (Coleoptera: Carabidae) contain resilin-rich structures.” Arthropod structure & development 49 (2019): 19-25.
Rossini, Carmen, Athula B. Attygalle, ANDREs GONZAlez, Scott R. Smedley, Maria Eisner, Jerrold Meinwald, and Thomas Eisner. “Defensive production of formic acid (80%) by a carabid beetle (Galerita lecontei).” Proceedings of the National Academy of Sciences 94, no. 13 (1997): 6792-6797.
Will KW, Attygalle A & Herath K. 2000. New defensive chemical data for ground beetles (Coleoptera: Carabidae): interpretations in a phylogenetic framework. Biological Journal of the Linnean Society 71, 459-481.
Will KW, Gill AS, Lee H, Attygalle AB. 2010. Quantification and evidence for mechanically metered release of pygidial secretions in formic acid-producing carabid beetles. Journal of Insect Science 10, available online: insectsicence.org/10.12.
Cover image: Bombardier Beetle – Paussinae subfamily, Gorongosa National Park, Mozambique. Photo: Judy Gallager (CC By 2.0)
Editing provided by Michael Callen

You say: “The 500 species are not all placed in the same formal taxonomic group but rather are found in four different subfamilies called tribes (Brachinini, Paussini, Ozaenini, or Metriini) of the ground beetle family Carabidae which contains more than 36,000 species. Could the Carabidae be a created kind? If this is so, the species that possess bombardier capabilities are but a small subset of this group and suggest that this specialized defensive capability evolved from ancestors that didn’t possess this specific defensive system.”
The information for the explosive chambers may have been cryptically present in the originally created kind. It only had to be derepressed through one single genomic macromutation to release this system as one program through epigenetic control. As you may know, transposons and genetic contexts heavily afect the epigenetic regulatory marks (methylation patterns of the DNA) can be regarded as VIGEs (variation and speciation-inducing genetic elemenst). If you can find remnant of this program in other members of the Carabidae family, this would compelling evidence the program was cryptically present in the Carbidae. In my book Darwin Revisted, which you may have read, I described several examples that prove this frontloading hypothesis.
Since you never refer to my papers in the JoC, which summarize these ideas, and which are often cited by the creation science community, I regard your contributions as biased. For your info I have again included my papers. Read them.
Best,
PB
Click to access Genetic_Redundancy.pdf
Click to access j22_3_68-76.pdf
Click to access j23_1_99-106.pdf
Click to access j23_1_107-114.pdf
LikeLike
The other 3 papers disappeared. They can be found here:
https://creation.com/images/pdfs/tj/j22_3/j22_3_68-76.pdf
https://creation.com/images/pdfs/tj/j23_1/j23_1_99-106.pdf
https://creation.com/images/pdfs/tj/j23_1/j23_1_107-114.pdf
LikeLike
What you are arguing seems like a fundamental revolution in biology, contrary to everything known and discovered from the last 200 years of work. Why don’t you write it and submit it to Nature instead of these self-publishing exercises? Then the whole scientific community can start benefiting from your work.
LikeLike
As there was no death prior to the fall, all defense and attack characteristics must have evolved afterwards. i.e. spikes,armor, venom and sprays.
LikeLike
Mati wrote: “As there was no death prior to the fall, all defense and attack characteristics must have evolved afterwards. i.e. spikes,armor, venom and sprays.” I trust you mean “According to YECS…” since the “no physical death” before the Fall doctrine is both Biblically unfounded and scientifically bankrupt. Besides what you mention, the many organisms that display elaborate camouflage and mimicry would make no sense except as means to elude predators, implying the existence of predators and physical death. One can’t say the prey bodies were just “modified” at the Fall, since some (like walking sticks, or walking leaf insets) would have to be entirely recreated, not just tweeked. Moreover, without physical death, ecosystems could not be sustained, and the earth would soon be awash in exponentially exploding populations of countless organisms, and the accompanying horrors. Strangely, YECs seldom deal with these serious problems, except in the most superficial and dismissive way. See:
http://paleo.cc/ce/nodeath.htm
https://biologos.org/common-questions/did-death-occur-before-the-fall
LikeLike
When I was a kid, I remember having a creationist book with a chapter on the bombardier beetle. I wish I could remember what book that was.
LikeLike
Stories of these beetles have around since the late 70s in the YEC literature. What is remarkable is how little that story has changed in 40 years despite how much more we know now. I was shocked–though not shocked either–how much more we know about these beetles than my hearing and reading YEC sources had told me.
LikeLike
What has changed, dear Joell? If there is now a gradual, chance driven evolutionary explanation for these wonderfully generated explosions, please let us know. IMHO, of much greater concern is that evolutionary theory has not changed for over 150 years. We are still told that selection of random mutation made everything. You and I know that to be untrue.
LikeLike
How can you lie like that? Aren’t you a Christian?
LikeLike
Peter, the main mechanism as you know is called natural selection. As as you also know, or should, it’s not merely “chance driven”, so if you are demanding a “chance driven” mechanism, you’re asking a misleading question or at best leading question. Ironically, it’s your theory that seems to be lacking a viable mechanism for the dramatic and quick changes you need to explain. You talk about TEs and such, but have never answered what specifically triggers or activates the genetic changes or expressions to match new or chancing environmental conditions. You suggested in the past that it was covered in your book, which many months ago you offered to send me, but never did. Recently you said it was because you did not have my email address, even though it is easy to find on the web, and I believe I left it in one of Joel’s posts early one. I’ve since left if for you in the Comments of another article by Joel that you were participating in, and will do so again here, in case you didn’t notice those. gkpaleo@yahoo.com
So if you were serious about your book offer, can you please contact me at that address? Thanks.
LikeLike
Duane Gish was fond of the example, mentioning it in several of his works (Dinosaurs by Design especially, where it was used as justification for dinosaurs being fire-breathing dragons).
LikeLiked by 2 people
Now that you say that, I do recall that the book explicitly used the bombardier beetle example to argue that certain dinosaurs with cavities in their skulls were the original “dragons” that used chemical excretions to expel flame or hot gas. However, that title doesn’t ring a bell.
LikeLike
I go into the grisly details of it in the “Dinomania” chapter of my old TIP work at http://www.tortucan.wordpress.com. In Gish’s case, the flame-throwing parasaurolophus was untenable at the skull level, as I explained, but it was really only a stage prop to allow him to bring up the beetle, and managed to get the biology of that wrong too.
Gish belongs to the old print era of YEC, and is less widely drawn on by today’s Young (Deranged) Turks (non Harun Yahya edition) who draw primarily on resources available online. That puts AiG apologetics as most commonly invoked (with their Creation.com divorced sibling comparable), but many are video YouTube addicts, and in that context Gish is almost forgotten. Given that dynamic, Joel’s contribution here is most relevant (and Jackson Wheat and I will definitely be including it in the second volume of “The Rocks Were There”).
LikeLike
You don’t understand. Even if all the different varieties of bombardier beetle belong to different kinds that were distinct at the time of the Flood, nonetheless all of those kinds could have been created with the genetic ability to give rise to bombardier beetles in some, though clearly not all, of their descendants. With God, all things are possible. With Young Earth creationism, no things are refutable.
LikeLiked by 1 person
Paul, while your comment that with YECs “no things are refutable” may be true in the sense that they have excuses and rationalizations for anything that challenges their view, in many cases (as with these beetles) salvaging a YEC position requires so much special pleading, neglect of or denial of important data, and sometimes demonstrable falsehoods, as to make their arguments thoroughly refuted, at least in my book. Moreover, it’s there are so many powerful lines of biologic, geologic, and paleontolgic evidence contradicting YECism that I consider it not only well refuted, but as thoroughly debunked as Flat Earthism. AIG’s current push for post-Flood hyper-rapid evolution is rich, since it contradicts not only lots of scientific evidence, but also so flatly contradicts many oft-repeated claims of past creationists, including leaders such as H Morris and Duane Gish.
LikeLike
Glen, I hope it’s clear that I am not defending the creationist position but, on the contrary, am arguing that it moves itself beyond science in two ways, by its acceptance of absurdly convoluted improbabilities, and by the fact that in so doing It is immunising itself against refutation
LikeLike
Paul, yes, I know you are no supporter of YECism, and did not mean to imply otherwise. I was just clarifying (for any new visitors here) that whereas YECs act like their view is impossible to refute in theory or practice, in fact it can and has been refuted in countless ways.
LikeLiked by 1 person
@peer terborg. You say: “The information for the explosive chambers may have been cryptically present in the originally created kind. It only had to be derepressed through one single genomic macromutation.”
It appears that you are making a prediction! Just like science does. But scientists modify their views, if their predictions are falsified. Are you?
LikeLike
Yes, it is a prediction. In my book written between 2003 and 2007 and published in dutch in 2009. The english version is stille available on Amazon:
https://www.amazon.de/Darwin-Revisited-understand-biology-century/dp/6202315113
It contains several daring predictions. In the meantime some of them have been born out as fact. When they are wrong, I’d simply admit that. Thus far there was no need to do so. The genomes are full of genes-in-waiting, I predicted, which simply can be activated by reshuffling of the genetic context, through the activity of TEs. That is what I wrote in 2006 and it is now beyond any doubt. So, genomes are frontloaded and “evolution” is an intrinsic characterstic of those genomes (which I coined Baranomes, i.e. fronloaded cryptic information containing multipurpose genomes).
LikeLike
I just read the description on the Scholar’s Press Website of the editorial process there. I couldn’t find any mention of peer-view. in fact it didn’t seem that much different from self-publishing with Amazon.
LikeLike
Peer/Peter wrote: “…genomes are frontloaded and “evolution” is an intrinsic characterstic of those genomes (which I coined Baranomes, i.e. frontloaded cryptic information containing multipurpose genomes). Two major problems. 1. Any “frontloading” would have been largely lost in the horrendously severe genetic bottlenecks at the time of the Flood, and 2. As far as I can see, you have no viable mechanism to explain adaptation, that is: what triggers the activation of specific TEs as an organism’s environment changes. You clearly need such triggers, especially if you reject natural selection. Can you summarize your supposed mechanism here, and how front-loading did not get largely wiped out during the Flood, even if these are covered in your book (are they?). Are you still willing to send me your book as you offered months ago? Thanks.
LikeLike
Reblogged this on Peddling and Scaling God and Darwin and commented:
For 40 years Creationists have told us about how the explosvie beetle – Bombardier – would have exploded if it evolved. It was always a bad argument, but those bad ideas never die.
It was a favourite with Gish in his gallops, but here it is exploded by Joel Duff
LikeLike
No it is not exploded by Joell Duff. Joell Duff forgot the review and include my papers on frontloading and baranomes. By selective reading and exclusion of my papers, he joins the group of theistic christians in my country, The Netherlands, which severly oppose me since 2009.
Joell, you will find the genes and construction-plan responsible for the explosion mechanism as remnants in the other subfamlies of Carabid beetles. This is a prediction which will come true.
LikeLike
You papers are not peer reviewed. They have nothing to contribute to scientific discussion. Don’t Christians count pride as a sin?
LikeLike
How many beetle baramins are there, Peter? And when were they allegedly created? Can your creationist genetics please specify the genomes of said created progenitors, to match the specificity which modern paleogenomics can regarding recent organisms? And if you accept a biblical global Flood, how did the beetle baramins manage through all that? What post-Flood speciation events (and global biogeographical shifts) occurred among them? Do please APPLY your otherwise meaningless and generic creationism to these issues, so they might crawl up to the status of at least a tentative model.
LikeLike
Peter, you seem to be missing the point about what Joel “exploded.” For years Gish and other YECs rejected and even mocked the idea that bombardier beetles evolved; now you and other YECs are implying that numerous bombardier species did evolve (from some unidentified original baramin), despite your rejecting natural selection as a mechanism, and with very little time to accomplish the astoundingly wide and rapid radiation. And again, what is your mechanisms is to activate specific genes or TEs to match environmental changes? What is your view on the age of the earth, and the date of Creation and the Flood? These questions are quite relevant to your claims.
LikeLike
Creationists having been sudyimg this beetle for forty years? Well first creation week would mean beetle(S) not made to kill or need to defend. its very suspect what pre fall insects were like or for. After the fall bodyplans changing indeed is becoming more hip in YEC. Rightly. So what a kind is in a beetles is not even close to figured out or the laws of it. Novel functions must be able to appear. my favorite is the tuatara, as i understand, which has a eye ish on the top of its head. never did this creature have this on creation week or even exist as this creature. so even a “eye” is able to be created out of existing dna .
LikeLike
Robert, just an FYI: given your track record of contributing nothing of substance to any discussion to which you have been a party, and the presence in this thread of more notable technically minded creationist apologists than you (Peer & Andy), might I suggest just sitting back and following the exchanges, rather than demonstrating yet again why you have the reputation you do.
LikeLike
Robert, as usual your rambling comments make little sense and thus not worth much response. However, for those wondering about your comment that the Tuatara has a third eye… The structure in question, sometimes called a pineal eye or parietal eye, is not an eye in the normal sense, but a light-sensitive region under the skin of some lizards, fish, and juvenile Tuataras (lost in adults), thought to be related to endocrine system development and perhaps other functions. In any case, there is no evidence that this feature came into existence at the “Fall”.
LikeLike
Do you have an answer to what Peer Terborg said about the origin of the Bombardier-Beetle (“The information for the explosive chambers may have been cryptically present in the originally created kind.”) and generally about your contributions (“Since you never refer to my papers in the JoC, which summarize these ideas, and which are often cited by the creation science community, I regard your contributions as biased. “)?
Thank you.
Warm greetings from Germany,
Miguel Goncalves
LikeLike
Fri 17th July 2020
Joel – concerning your paragraph: “The 500 species are not all placed in the same formal taxonomic group but rather are found in four different subfamilies called tribes (Brachinini, Paussini, Ozaenini, or Metriini) of the ground beetle family Carabidae which contains more than 36,000 species. Could the Carabidae be a created kind? If this is so, the species that possess bombardier capabilities are but a small subset of this group and suggest that this specialized defensive capability evolved from ancestors that didn’t possess this specific defensive system.”
As Peer Terborg has written above you are making a leap in your assumptions in your supposed critique of the Creation position as regards the origin of the blasting apparatus of the Bombardier Beetle. None of us can be sure of the ancestral Carabidae beetle genetic material, but it is evident by observation today (and we all agree with you) that “the species that possess bombardier capabilities are but a small subset of this group [of Carabidae].” But you then make a serious non-sequitur by stating that “this specialized defensive capability evolved from ancestors that didn’t possess this specific defensive system.” On what authority are you making such a sweeping statement? The actual scientific evidence indicates another much more plausible explanation. The information and engineering necessary for making an irreducibly complex blasting apparatus (which uses many interconnected and interwoven systems) is not obtained by a linear trial and error route from DNA which did not have such information at the outset. The recent advances in the engineering understanding of the bombardier beetle defence aparatus has shown the sophistication of multi-layered interacting systems which include
(a) an inlet chemistry tube which is of the order of microns in diameter where reactants Hydrogen Peroxide and Hydroquinone are produced but are inhibited from reacting.
(b) a catalyst production system – the catalysts Peroxidase and Catalase which are in the combustion chamber and are probably in crystalline form
(c) the inlet pinching valve system and the exhaust pressure-relief valve system
(d) the controlled explosive chemistry system within a combustion chamber made from low thermal conductive cuticle to contain the initial temperature and pressure rise
(e) a special sensory mechanism for obtaining the direction from which the predator is attacking from
(f) an actuator system taking information from the above sensor to bring the exhaust turret into the correct position to bring the blast onto the attacker
(g) a system to replenish and draw on the reactants to support a stream of explosive blasts emitted at 400-500 times per second for approximately 2-3 seconds
There are many other interlocking systems, and an exhaustive list would also include the genetic interrelated software control systems which will be found by considering the genome of the carabidae beetles.
In the work we have done at Leeds and other academic institutions in the last 15 years in biomimetics, we have copied only the valve system of the beetle and the sophistication involved in this biomimetic challenge on its own, took careful research and design to achieve just one part of this integrated system in the beetle. This work is described at https://digitalcommons.cedarville.edu/icc_proceedings/vol8/iss1/6/ To suggest that the complete set of integrated chemical / mechanical / fluid mechanical / biochemical interacting systems of the bombardier beetle could emerge through the neo Darwinian synthesis of mutations and Natural Selection is frankly not backed by careful experimental observations.
Though none of us can go back in time and observe the original ancestral Carabidae of the past, it is perfectly feasible that the present variety is a combination of three main effects :
(1) alleles of genetic material providing the variety of genetic expression from an original rich pool in the ancestral kind.
(2) natural selection preferring some descendants with particular attributes which were better suited to particular different geographical locations – that of course is not evolution where entirely new traits are made which were not coded for previously; rather this is just simply variation and selected genetic expression. The recent advance of epigenetic research is also now showing that environmental pressures can switch genes on or off which will further enhance such variations. Peer Terborg is making this same point when he states “The information for the explosive chambers may have been cryptically present in the originally created kind. It only had to be derepressed through one single genomic macromutation to release this system as one program through epigenetic control.”
(3) mutations will lead to loss of genetic information by corruption of coding strands within some of the chromosomes. It is my conviction (though this has yet to be be proved) that today the many carabidae which do not blast but simply send out a mixture of vapour and small droplets of quinones, are in fact mutant descendants which have lost the blasting capability of the ancestral carabidae, rather than such a system emerging by small evolutionary steps.
For those interested my recent ICC 2018 paper available is at https://digitalcommons.cedarville.edu/icc_proceedings/vol8/iss1/6/
Professor Andy McIntosh
Professor Andy C. McIntosh DSc, FIMA, C.Math, FEI, C.Eng, FInstP, MIGEM, FRAeS
Emeritus Professor of Thermodynamics, School of Chemical and Process Engineering, University of Leeds, LEEDS, UK
Adjunct Professor, Department of Agricultural and Biological Engineering. Mississippi State University, Starkville, Mississippi, USA
https://engineering.leeds.ac.uk/staff/46/Professor_Andy_McIntosh
https://professorandymcintosh.academia.edu
http://bombardierbeetle.org
LikeLike
Thanks for the comment. Hectic day at the moment but have printed paper which I hadn’t read so thanks, and will read this evening. Definitely some good stuff to consider here. Had similar feedback elsewhere that I will address in an addendum at some point hopefully soon.
LikeLike
Andy, You seem to imply that you and Peer largely agree in your views, and yet you seem to imply that natural selection had a major role (acting on supposedly “frontloaded” varation), while Peer seems to argue that natural selection has little if any role in evolution. Have I misunderstood either of you? And as I’ve often asked Peer, how is the “frontloaded” variation not goiing to be largely lost during the severe bottlenecks at the time of the flood. Or are you suggesting that God “reloaded” the genes?
LikeLike
Andy, you wrote: “The recent advance of epigenetic research is also now showing that environmental pressures can switch genes on or off which will further enhance such variations.” First, you and peer seem to ignore the possibility that to the extent such switches exist, they themselves could have evolved. Second, if you are suggesting that all such switches (and supposed vatiations they act on) were present in the genes of originally created organisms’, how was most of these variations not lost the severe bottleneck at the time of the Flood? Third, how would such switches “further enhance such variations.” as you claim? Do you not accept that natural selection acting on mutations can produce adaptations? Otherwise, how are thousands of new alleles going to be produced since the Flood? After all, in general a pair of animals can only have four alleles for a specific gene, whereas many animal species today have dozens or more alleles for specific traits, or affecting those traits. That implies that lots of new genetic information has been produced through evolution, despite YEC claims to the contrary. Said another way, you can’t have pre-designed switches acting on lots of variation if most of the variations were lost at the Flood.
Last, what is your view on the age of the Earth? Do you agree with ICR’s admission in their RATE project report that the Earth’s crust records far too much radioactive decay to fit a YEC timetable? Unable to face the logical conclusion that their view was wrong, they proposed ad-hoc miracles (to vastly accelerat nuclear decay), even though it would serve no purpose other than to fry the Earth, make it look old, require more miracles to counteract. And after all that, it still does not explain the observed sloping pattern of radio dates by several methods. Do you know of any remotely reliable dating method that points to an earth of only several thousand years old?
LikeLike
Just to be clear, based on design characteristics, how many designers do you think were involved in all this, Andy? We know, for example, that while the indisputably designed Cadillacs and Lincolns shared many common features (wheels, doors, etc), they were not designed by the same designers, nor only as individuals, but rather teams (John Najar, for example, leading the team that produced the 1958 Lincoln). So how many did you have in mind here? Certainly not the same designer(s) that made dinosaurs, right? Or therapsids (those designers really adored evolution, though, didn’t they, littering the fossil record with specific examples precisely matching evolutionary expectations, such as the dual-jawed probainognathids). And are any of your proposed designers currently active in the field, or are they presently retired or extinct? Just as the designer of the original Thunderbird is no longer working at Ford, on account of not being alive.
And when exactly was this purported design event (or events) with the original bombardier beetles? You left out chronology in your summary. Was this an intentional oversight? After all, if a proposed model is aiming to be taken seriously as a competitive alternative scientifically, such details should be made clear, so their viability may be properly assessed, openly and honestly.
LikeLike