Nearly two decades ago I published an article in Perspectives on Science and Christian Faith titled “Flood Geology’s Abominable Mystery.”(Duff 2008) The title borrowed from Darwin, who famously called the sudden appearance of flowering plants in the fossil record an abominable mystery, but I applied the phrase to a different problem. If a global Flood had laid down the world’s sedimentary rocks in a single year, as young-earth creationists maintain, then the distribution of plant fossils within those rocks should look nothing like what we actually observe. In particular, the distribution of microscopic pollen and spores should not exhibit the orderly, globally consistent succession that palynologists have documented at thousands of sites on every continent.

That was 2008. In the years since, the problem has only sharpened. Hundreds of additional palynological studies have confirmed the pattern. Radiometric dating of volcanic ash layers within coal seams has become more precise. The biomass calculations that constrain pre-Flood scenarios have become more refined. And to its credit, the YEC community has produced more sophisticated engagement with the plant fossil record than it had two decades ago. That includes a remarkable internal dispute in which two of YEC’s leading researchers publicly called for abandoning the dominant model of Carboniferous coal formation.
Despite all this, the plant fossil record remains among the least-discussed challenges to flood geology in popular-level Christian apologetics. That seems worth correcting. My purpose in this post is not to ridicule YEC authors many of whom are sincere believers doing the best science they know how to do, but to survey the full range of models they have proposed for the plant fossil record, present each one fairly, and then ask how well each holds up when tested against what the rocks actually contain. I write as a confessional Reformed Christian and a professional plant systematist. I have a dog in both fights, and I want both done well.
The Ordered Succession Everyone Now Admits
Let’s start with what is no longer in dispute. Plant fossils appear in the geological column in an ordered sequence that is remarkably consistent worldwide. Non-vascular plants such as the mosses, hornworts and liverworts (the bryophytes) appear earliest. Spore-bearing vascular plants — the lycopsids, sphenopsids, and ferns — dominate the lower Paleozoic and especially the Carboniferous. Gymnosperms, the seed-bearing but flowerless plants (pines, firs, cycads, ginkgos), appear in the mid-Paleozoic and become abundant in the Mesozoic. Flowering plants (the angiosperms) do not appear until the early Cretaceous, and the grasses and the sunflower family do not appear until even later. The pattern is not subtle, and it is not the product of evolutionary bias in sample selection.
That this pattern is real, and not an artifact of misclassification or circular reasoning, has been conceded by some of the most prominent voices within YEC scholarship. In a 2002 exchange in the Journal of Creation, John Baumgardner pushed back against Michael Oard’s attempt to deny the reality of fossil succession. Baumgardner wrote that “fossil succession represents an undeniable reality of what creationists and evolutionists alike observe in the rock strata” and warned that creationists who deny it cannot make “genuine progress in reconstructing the actual history of the Earth in light of God’s revelation.” Kurt Wise, in his 2003 ICC paper on the floating forest hypothesis, went further. He wrote that the order of higher plant first appearances is “highly correlative” with the order of evolutionary branching predicted from cladograms, and that “the probability that this pattern could be arrived at randomly is so low as to suggest that an explanation is required in the young-age creation model.” That is a remarkable admission from a Harvard-trained paleontologist committed to young-age creation.
The palynological pattern is the same, only more precise. Microscopic spores and pollen, which are abundantly preserved and easily identified to broad taxonomic groups, follow the macrofossil order with high fidelity. Spores from non-vascular and early vascular plants are found in the lowest plant-bearing layers. Gymnosperm pollen appears later. Angiosperm pollen (flowering plant pollen) appears no earlier than the early Cretaceous (less than 140 million years ago), despite extensive sampling on every continent.
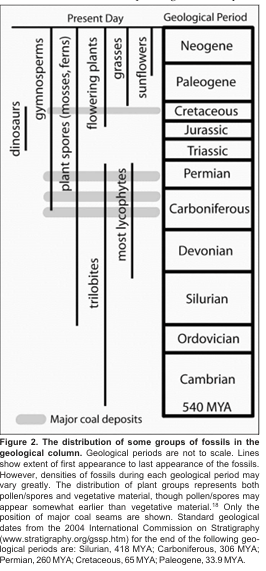
The coal record is a particular sub-challenge that deserves its own attention. The world’s great Carboniferous coal deposits — billions of tons of fossil plant material across Europe, North America, China, and Australia — contain exclusively lycopsid, sphenopsid, and fern debris along with primitive gymnosperm pollen. Not a single trace of angiosperm wood, leaf, flower, or pollen has been documented in any Carboniferous coal seam anywhere on Earth.
This is the pattern every YEC model must explain. Let’s see what they have proposed.
The Roster of YEC Models: A Charitable Survey
Readers accustomed to thinking of “the YEC position” on plants may be surprised to discover that there is not one. YEC paleobotany is a family of competing proposals, some of which actively contradict each other, and which collectively have produced no agreed-upon model of how a single year-long global Flood deposited the plant fossil record we actually observe. Let me outline the major proposals, in roughly the order they emerged.
The Classic Trio: Ecological Zonation, Hydrodynamic Sorting, and Differential Escape
The earliest and still most popular YEC explanation for fossil succession was developed by Whitcomb and Morris in The Genesis Flood (1961), drawing on Harold Clark’s The New Diluvialism (1946) and elaborated in Henry Morris’s Scientific Creationism (1974). It combines three mechanisms.
Ecological zonation proposes that pre-Flood biomes occupied different elevations on the pre-Flood landscape, and that as floodwaters rose they buried these biomes in elevational sequence. Marine creatures and shoreline organisms first, then lowland terrestrial plants and animals, then upland flora and fauna at the top. Hydrodynamic sorting adds that within each pulse of sediment, particles of similar size, shape, and density settle together: denser and more spheroid organisms sink lower, more buoyant ones ride higher. Differential escape adds the further factor that mobile organisms (animals, especially mammals) could flee to higher ground, while plants, lacking mobility, were buried where they grew.
Taken together, this trio has some initial plausibility. It explains the rough ordering of marine versus terrestrial fossils, and it provides a framework that lay readers can visualize. It is the model most YEC popular literature still presents, and it appears in some form on signage at the Ark Encounter and the Creation Museum.
The Floating Forest / Pre-Flood Biome Hypothesis
The most imaginative and theoretically ambitious YEC plant model was developed in stages by Joachim Scheven in the Creation Ex Nihilo Technical Journal (1996) and, most fully, by Kurt Wise in the Proceedings of the Fifth International Conference on Creationism (2003). The core idea is that the arborescent lycopsids of the Carboniferous — Lepidodendron, Sigillaria, Lepidophloios, and their relatives — were not terrestrial swamp trees. They were, on this view, the dominant inhabitants of a continent-scale floating biome on the surface of the pre-Flood ocean.
The case for this interpretation rests on the genuinely unusual anatomy of these extinct plants. Their trunks were not solid wood but rather a thick outer cortex surrounding a relatively small vascular strand, which floating-forest advocates interpret as an adaptation for buoyancy. Their root systems — the genus Stigmaria — branched horizontally rather than penetrating downward, with radiating rootlets that floating-forest advocates compare to bottle brushes designed to interlock with neighbors to form a stable mat. Their reproductive structures are interpreted as aquacarps, propagules designed to float and germinate on water.
On this model, the Flood destroyed the floating biome from the outside inward. The ecological zonation within the mat which includes open-water edge species first, more interior species later, generated the observed stratigraphic order of Carboniferous plant fossils. Answers in Genesis has popularized this model in its publications and exhibits.
The Log Mat / Allochthonous Coal Model
Distinct from the floating forest but often conflated with it is Steven Austin’s allochthonous coal model, developed in his 1979 Penn State dissertation on the Kentucky No. 12 coal bed and elaborated in his ICR Impact pamphlet Mt. St. Helens and Catastrophism (1986). After the 1980 eruption of Mount St. Helens, Austin and Harold Coffin observed that Douglas firs swept into Spirit Lake formed a floating log mat, and that bark abraded from these logs settled to the lake bottom as a thick organic deposit. Austin extrapolated this real-time analog to the Carboniferous: most coal, on his model, derives from water-transported vegetation ripped up by floodwaters, with the dominance of bark-rich coal lithotypes explained as a natural consequence of mechanical abrasion in a turbulent log raft.
Austin’s model is distinct from Wise’s because it does not require a living, ecologically structured floating biome. It only requires catastrophic transport of pre-Flood terrestrial vegetation. Austin and Sanders restated this model in 2018 in the proceedings of the Eighth ICC.
Tectonically-Associated Biological Provinces
John Woodmorappe, writing under that pseudonym in the Creation Research Society Quarterly (1983) and in Studies in Flood Geology (1999), proposed a different model entirely. On Woodmorappe’s view, different pre-Flood biomes occupied different paleogeographic and tectonic zones. These are distinct biogeographic provinces with distinct floras and faunas. As the Flood progressed, these zones were buried in sequence as tectonic activity spread across the pre-Flood Earth. Distinct plant provinces in the fossil record thus reflect distinct pre-Flood geographic biomes, not deep time.
Woodmorappe’s TABs model is the only YEC proposal that even attempts to address the paleobiogeographic problem — why, for instance, the extant Glossopteris flora is restricted to the now-separated southern continents and entirely absent from contemporaneous equatorial deposits.
Post-Flood Rapid Recolonization
Finally, YEC scholars differ on where to place the Flood/post-Flood boundary, but most recent proposals place it at the Cretaceous-Paleogene boundary or somewhere in the Cenozoic. Plants found above this boundary are explained as survivors that rapidly recolonized the barren post-Flood landscape — from seeds preserved in floating debris, from vegetative fragments, or from rapid diversification within originally created “kinds” or baramins. Roger Sanders’s 2013 ICC paper is the most serious YEC engagement with angiosperm family-level diversification; he proposes that different angiosperm families show varying ratios of “originally designed diversity” versus post-Flood speciation — what I have elsewhere called the hyper-speciation paradox, in which YEC models require evolutionary rates that outstrip anything mainstream biology has ever proposed.
When YEC Scholars Disagree With Each Other
The most productive critiques of any research program come from within. So before turning to mainstream paleobotanical evidence, it is worth noting that YEC paleobotany is currently in the middle of a public, unresolved internal dispute that cannot be dismissed as the work of hostile outsiders.
In 2016, Tim Clarey and Jeffrey Tomkins, who are both senior researchers at the Institute for Creation Research, the oldest of the major YEC organizations, published a paper in the Creation Research Society Quarterly titled “An Investigation into an In Situ Lycopod Forest Site and Structural Anatomy Invalidates the Floating-Forest Hypothesis.” A year later they followed with an ICR Article titled, with rather striking directness, “Sinking the Floating Forest Hypothesis.” Their findings are worth describing carefully. Examination of in-situ lycopod sites (particularly Fossil Grove in Glasgow) and careful anatomical study of preserved lycopsid trunks led them to conclude that lycopsid trunks were not hollow, that Stigmaria root systems penetrated soils in growth position, and that the floating-forest model is geologically and anatomically untenable. They explicitly recommended that the YEC community abandon it.
Wise responded in Answers Research Journal in 2018, defending the floating biome interpretation of Fossil Grove and disputing the anatomical conclusions. The debate remains unresolved. As of 2026, the YEC community has no agreed-upon model of how Carboniferous coal beds and their associated plant fossils were laid down during the Flood year. The two most-developed proposals — the floating forest and the allochthonous log mat — are not just unverified; they are being publicly contested by leading YEC researchers in YEC journals.
Then there is the biomass problem, which is acknowledged obliquely in the YEC literature but never squarely addressed. Cleal and Thomas (2005) estimated that peak Carboniferous coal-forming biomass sequestered carbon at 13 to 47 billion tonnes per year over a roughly 1.2 million square kilometer area of equatorial Pangaea. The total mass of carbon now locked in Carboniferous coal worldwide is staggering — multiple orders of magnitude greater than the entire modern global plant biomass, which Bar-On and colleagues (2018) estimated at about 450 gigatonnes of carbon. On a conservative estimate, the pre-Flood biosphere would need to have exceeded modern plant biomass by something between five and twenty times in order to supply the world’s known Carboniferous coal deposits alone.
The floating-mat model does not solve this problem; it intensifies it, because a floating biome adds a constraint (where exactly did all that biomass live, and how was it ecologically sustained?) without adding any new carbon source. If most of the ocean surface where covered with vast floating forests how did light get to photosynthetic phytoplankton in the water which are the primary producers providing energy for aquatic systems?
And then there is Joggins. The Joggins Formation in Nova Scotia is the classic Carboniferous coal-bearing succession on Earth, a UNESCO World Heritage site. Calder and colleagues (2006) and Davies and Gibling (2003) have documented more than 60 stacked horizons of rooted lycopsid forests through 1,425 meters of section. Each forest sits atop its own paleosol. Each was buried in place by sediment delivered during salt-withdrawal-driven subsidence. This is exactly what the floating-mat model cannot produce — and it is the very feature, polystrate trees, that YEC popular literature has long pointed to as supposed evidence against deep time. The Joggins succession is not a single buried forest; it is sixty buried forests stacked on top of each other, each rooted in soil. No model on the YEC side accounts for this.
The Problem That Size Cannot Solve
Set aside, for a moment, every macrofossil. Set aside the lycopsid trunks, the Joggins paleosols, the carbon mass. Focus on something much smaller: pollen and spores.
Pollen and spores are produced in astonishing quantities by living plants. A single pine cone can produce 600,000 pollen grains. A mature pine tree produces around 350 million pollen grains per year. An oak tree produces over 100 million. Pollen production in a typical temperate forest is several billion grains per hectare per year. These particles are tiny (typically 10 to 50 micrometers in size) and their resistant outer walls make them extraordinarily durable. They are found in modern oceans thousands of miles from any land, in Greenland ice cores, in mammal feces, in the digestive tracts of preserved insects, and in lake sediments going back through the entirety of the Quaternary.
If pollen-producing plants were present in the pre-Flood world, and they must have been, given the explicit Genesis references to fruit-bearing trees in Eden, Abel’s grain offering, the gopher wood of the Ark, the olive leaf brought back by the dove, and Noah’s post-Flood vineyard, then their pollen should be present somewhere in the lower portions of the Flood-deposited rock record. It is not.
Here is the heart of the problem and the focus of my paper Flood Geology’s Abominable Mystery (Duff 2008), and it is one that no proposed YEC sorting mechanism can plausibly address. Gymnosperm pollen and lycopsid spores are virtually identical in size, density, and shape. Both fall in the 10 to 50 micrometer range. Both have resistant outer walls. Both float similarly in water and disperse similarly in air. No known hydrological sorting mechanism — laboratory or field, modern or modeled — can separate them on a global scale. The careful taphonomic studies of modern pollen transport in rivers, estuaries, and ocean currents (Moss et al. 2005; Chmura and Eisma 1995; Brush and Brush 1972) show that even local sorting is rough, partial, and incomplete. A globally turbulent Flood would produce less sorting, not more.
And yet what the fossil record shows is exquisitely clean global sorting. Spores in the lower Paleozoic. Gymnosperm pollen in the upper Paleozoic and Mesozoic. Angiosperm pollen only from the Cretaceous upward. Grass pollen whch is distinctive, easily identified, and produced in enormous quantities by a worldwide modern flora, appears no earlier than the late Cretaceous. Asteraceae pollen, from the second-largest family of flowering plants, does not appear until the late Paleocene.
Since 2008, the palynological record has only become more finely resolved. Hundreds of additional studies have confirmed the pattern. The absence of angiosperm pollen from Paleozoic strata is not a data gap but rather it is a positive finding, repeatedly tested at sites on every continent.
The Most Recent YEC Attempt to Solve the Problem
The most recent serious YEC engagement with this problem is W.H. Johns’s 2022 paper in the Journal of Creation, “Puzzle of the missing angiosperms in the fossil record.” Johns attempts to reframe the absence of pre-Cretaceous angiosperm pollen as a problem for standard chronology rather than for Flood geology, and revisits the disputed “out-of-place pollen” claims from the Grand Canyon.
That history is worth reviewing. In 1966 and again in 1974, Clifford Burdick reported finding angiosperm pollen in the Precambrian Hakatai Shale of the Grand Canyon. If true, this would have placed flowering plants more than 500 million years before they appear in conventional stratigraphy. Arthur Chadwick, a young-earth creationist himself, conducted careful follow-up testing and concluded that Burdick’s findings were the result of modern pollen contamination of microscope slides (Origins 8, 1981). A later group of YEC researchers (Howe, Williams, Matzko, and Lammerts, in CRSQ, 1986–1988) attempted to vindicate Burdick’s original claims, but their contamination controls were inadequate, and the mainstream scientific community’s assessment has not changed.
John 2022 paper concludes that the “lowest three fourths of the fossil record have no valid evidence of angiosperm presence.” All previous reports of ancient angiosperm pollen he agrees are more likely the result of contamination. He suggests that angiosperm pollen being found throughout the geological column is “an argument that creationists should no longer use” and I very much agree with that conclusion.
What I want YEC readers to notice is what this episode actually shows. If a global Flood had truly mixed angiosperm pollen into Precambrian sediments, we would not need a single contested study from a single location to demonstrate the fact. We would expect to find angiosperm pollen routinely, at hundreds of sites, in every major palynological survey. The fact that, after sixty years of searching by both mainstream and YEC researchers, the case for pre-Cretaceous angiosperm pollen reduces to a handful of contested findings from a single canyon — and that even YEC researchers like Chadwick have rejected those findings — is itself powerful negative evidence.
On a conservative estimate, the plant fossil record has now been systematically sampled at thousands of locations worldwide, in basins on every continent, in formations spanning the entire Phanerozoic. The probability that the globally consistent absence of angiosperm pollen from the lower two-thirds of the geological column resulted from chance sorting during a single-year flood is not merely low. It approaches zero.
A World Too Organized to Have Been Mixed
Even if some sorting mechanism could be devised to separate spores from pollen which, as I have argued, none can, there is a further problem that compounds the difficulty. The Late Paleozoic plant fossil record is not just stratigraphically ordered; it is geographically structured.
Paleobotanists have long recognized four classical Late Paleozoic floral provinces, each diagnosable by both macrofossils and palynology. The Euramerican coal-swamp flora dominates equatorial Pangaea — the lycopsid forests of what is now eastern North America, Europe, and North Africa. The Gondwanan Glossopteris flora is restricted to high southern paleolatitudes and is found today across South America, Africa, India, Australia, and Antarctica. The Angara flora occupied present-day Siberia. The Cathaysian flora occupied what is now East Asia. These provinces are not interchangeable. They are characterized by distinct genera and species, and their boundaries grade into mixed-province assemblages precisely along documented plate-tectonic suture zones.
This is exactly what mainstream paleobiogeography predicts and exactly what a globally mixing flood cannot produce. Woodmorappe’s TABs model attempts to address this, but it provides no quantitative mechanism for how a year-long flood preserved such tight provincial fidelity across millions of square kilometers, especially for wind-dispersed pollen, which on any plausible model would have been mixed by global atmospheric circulation long before the Flood ended.
Consider the Glossopteris problem alone. This flora is found across five now-separated continents, all in what were Gondwanan paleolatitudes. It is entirely absent from contemporaneous equatorial Euramerican deposits. Ecological zonation which relies on elevation, not latitude, cannot do this work. A floating biome destroyed from the outside inward cannot do this work either. The geographic structure of the Late Paleozoic plant fossil record demands deep time and continental drift.
What a Good Model Would Need to Explain
Rather than simply cataloguing problems, let me try to sketch what a successful YEC model of the plant fossil record would actually need to accomplish.
Such a model would need to explain, at minimum: first, the global consistency of pollen and spore succession across unconnected basins on every continent; second, the biomass paradox that requires a pre-Flood plant biosphere five to twenty times larger than the modern one; third, the dozens of stacked rooted lycopsid forests at Joggins and similar sites, each in its own paleosol; fourth, the four geographically distinct Late Paleozoic floristic provinces and their suture-zone gradations; fifth, the complete absence of angiosperm pollen from Paleozoic strata worldwide despite extensive sampling; and sixth, the internal ecological coherence of Carboniferous coal assemblages, which are not randomly mixed but organized into recognizable plant communities with internally consistent water-table chemistry.
That is a substantial explanatory burden. The current state of YEC paleobotany with no unified model, two leading researchers publicly disputing the dominant hypothesis, and a biomass problem nobody has addressed, does not inspire confidence that such a model is forthcoming. The honest response, I would argue, is to take seriously the possibility that the plant fossil record means what mainstream paleobotany has long understood it to mean.
Blessings,
Joel
References
Scientific Literature
Bar-On, Y. M., Phillips, R., and Milo, R. 2018. The biomass distribution on Earth. Proceedings of the National Academy of Sciences 115(25): 6506–6511. DOI: 10.1073/pnas.1711842115.
Brush, G. S., and Brush, L. M. 1972. Transport of pollen in a sediment-laden channel: A laboratory study. American Journal of Science 272(4): 359–381. DOI: 10.2475/ajs.272.4.359.
Calder, J. H., Gibling, M. R., Scott, A. C., Davies, S. J., and Hebert, B. L. 2006. A fossil lycopsid forest succession in the classic Joggins section of Nova Scotia: Paleoecology of a disturbance-prone Pennsylvanian wetland. In Wetlands Through Time, Geological Society of America Special Paper 399, 169–195. DOI: 10.1130/2006.2399(09).
Chmura, G. L., and Eisma, D. 1995. A palynological study of surface and suspended sediment on a tidal flat: Implications for pollen transport and deposition in coastal waters. Marine Geology 128: 183–200.
Cleal, C. J., and Thomas, B. A. 2005. Palaeozoic tropical rainforests and their effect on global climates: Is the past the key to the present? Geobiology 3(1): 13–31. DOI: 10.1111/j.1472-4669.2005.00043.x.
Davies, S. J., and Gibling, M. R. 2003. Architecture of coastal and alluvial deposits in an extensional basin: The Carboniferous Joggins Formation of eastern Canada. Sedimentology 50: 415–439. DOI: 10.1046/j.1365-3091.2003.00553.x.
DiMichele, W. A., and Phillips, T. L. 1994. Paleobotanical and paleoecological constraints on models of peat formation in the Late Carboniferous of Euramerica. Palaeogeography, Palaeoclimatology, Palaeoecology 106(1–4): 39–90.
Duff, R. J. 2008. Flood geology’s abominable mystery. Perspectives on Science and Christian Faith 60(3): 166–177.
Gastaldo, R. A. 1984. A case against pelagochthony: The untenability of Carboniferous arborescent lycopod-dominated floating peat mats. In K. R. Walker, ed., The Evolution-Creation Controversy: Perspectives on Religion, Philosophy, Science and Education, Paleontological Society Special Publication 1, 97–116.
Gastaldo, R. A. 2009. Debates on autochthonous and allochthonous origin of coal: Empirical science versus the diluvialists. Paleontological Society Papers 15: 233–256.
Herendeen, P. S., Friis, E. M., Pedersen, K. R., and Crane, P. R. 2017. Palaeobotanical redux: Revisiting the age of the angiosperms. Nature Plants 3: 17015. DOI: 10.1038/nplants.2017.15.
Lyons, P. C., Krogh, T. E., Kwok, Y. Y., Davis, D. W., Outerbridge, W. F., and Evans, J. H. T. 2006. Radiometric ages of the Fire Clay tonstein [Pennsylvanian (Upper Carboniferous), Westphalian, Duckmantian]: A comparison of U-Pb zircon single-crystal ages and 40Ar/39Ar sanidine single-crystal plateau ages. International Journal of Coal Geology 67(4): 259–266.
Moss, P. T., Kershaw, A. P., and Grindrod, J. 2005. Pollen transport and deposition in riverine and marine environments within the humid tropics of northeastern Australia. Review of Palaeobotany and Palynology 134(1–2): 55–69. DOI: 10.1016/j.revpalbo.2004.11.003.
Young-Earth Creationist Sources Discussed
Austin, S. A. 1986. Mt. St. Helens and catastrophism. ICR Impact 157.
Austin, S. A., and Sanders, R. W. 2018. Historical survey of the floating mat model for the origin of Carboniferous coal beds. In J. H. Whitmore, ed., Proceedings of the Eighth International Conference on Creationism, 277–286. Pittsburgh, PA: Creation Science Fellowship.
Baumgardner, J. 2002. A constructive quest for truth. Journal of Creation 16(1): 79–81.
Burdick, C. L. 1966. Microflora of the Grand Canyon. Creation Research Society Quarterly 3(1): 38–50.
Burdick, C. L. 1974. More Precambrian pollen. Creation Research Society Quarterly 11: 122–123, 126.
Chadwick, A. V. 1981. Precambrian pollen in the Grand Canyon—a re-examination. Origins 8(1): 7–12.
Clarey, T. L., and Tomkins, J. P. 2016. An investigation into an in situ lycopod forest site and structural anatomy invalidates the floating-forest hypothesis. Creation Research Society Quarterly 53(2): 110–122.
Clarey, T. L., and Tomkins, J. P. 2017. Sinking the floating forest hypothesis. Acts & Facts 46(8). Institute for Creation Research. Published July 31, 2017.
Howe, G. F., Williams, E. L., Matzko, G. T., and Lammerts, W. E. 1988. Creation Research Society studies on Precambrian pollen—Part III: A pollen analysis of Hakatai Shale and other Grand Canyon rocks. Creation Research Society Quarterly 24(4): 173–182.
Johns, W. H. 2022. Puzzle of the missing angiosperms in the fossil record. Journal of Creation 36(2): 106–111.
Sanders, R. W. 2013. The fossil record of angiosperm families in relation to baraminology. In M. Horstemeyer, ed., Proceedings of the Seventh International Conference on Creationism. Pittsburgh, PA: Creation Science Fellowship.
Scheven, J. 1996. The Carboniferous floating forest—an extinct pre-Flood ecosystem. Creation Ex Nihilo Technical Journal 10(1): 70–81.

How did YEC ever cite polystrate trees as evidence in their favor?
LikeLike